Entomology and
Applied Science Letters
Applied Science Letters
 2023
Volume 10
Issue 4
2023
Volume 10
Issue 4

Biodiversity protection is crucial due to the impact of human activities on ecosystems. Anthropogenic activities, such as advancing agricultural frontier, deforestation, and the introduction of exotic species, are the main drivers of changes in biodiversity. Entomofauna, especially insects, play a vital role in ecosystem indicators of environmental quality and contribute to key functions such as pollination and decomposition. The present research focuses on the understory entomofauna in the Faculty of Veterinary Sciences of the National University of Rosario, Argentina. Several samplings were carried out in 2022 in five different microsites regarding herbaceous, shrub, and tree vegetation, using pitfall traps to capture arthropods. A total of 2631 individuals of 68 morphospecies belonging to five classes, 15 orders, and 43 families were captured. The class Insecta was the most diverse and abundant, accounting for 82.35% of the morphospecies and 35.80% of the total abundance. The class Malacostraca, although with only two species, was dominant in abundance (58.57%). Microsite diversity showed that microsites 5 and 1 had the highest biodiversity. The results showed low similarity between microsites, indicating the influence of the landscape on entomofaunal composition. This study provides an essential baseline on understory entomofauna in the Faculty of Veterinary Sciences. The diversity found highlights the importance of this habitat and the identification of key species such as Armadillidium vulgare and Enthomobryidae sp. to their role in ecosystem services. This research lays the groundwork for future comparative studies and highlights the need to consider biodiversity in conservation and environmental management politics agendas.
INTRODUCTION
Biodiversity protection is a relevant issue in global conservation. Given the environmental impact of human activities on natural systems, biodiversity knowledge is a worldwide challenge [1].
Anthropogenic activities such as overexploitation and habitat loss, deforestation, pollution, and the introduction of exotic species, among others, are the main forces driving changes in the diversity and composition of communities worldwide [2-4].
Entomofauna is the dominant group of animals on earth, with the number of known species exceeding three times that of all other animal species combined. These living beings conquered the planet more than 350 million years ago, and their importance in the world is such that without arthropods, life would be very different from how we know it, which is why they form an important component of ecosystem diversity [5].
Generally, the degree of disturbance of a local ecosystem is interpreted by the presence or absence of various arthropods, their variations in abundance, diversity, and composition of biological groups. Insects, for example, are excellent indicators frequently used to determine and monitor certain ecosystems' environmental quality [6]. The latter is explained by being one of the most diverse and widely distributed groups, rapid identification, simple sampling, convenient size, and predictable response to environmental variations, and being a useful, reliable, and economical tool [7]. In addition, entomofauna play important roles in pollination, seed dispersal, aeration, and soil rotation and act as parasites, decomposers of organic material, and nutrient recyclers. In addition, they form the basis of many food chains and ecological interactions, promote soil fertility and structure, and provide biological control mechanisms [8].
All of the above attributes cause arthropod communities to vary according to the degree of disturbance (natural and anthropogenic), abundance, diversity, and composition of biological groups, as they are strongly correlated with ecosystem functioning and reflect habitat heterogeneity, development, and recovery. Likewise, structural arthropod communities reflect the degree of fragmentation and isolation of ecosystems in the landscape [9].
In recent decades, in Argentina, the simplification of the landscape due to the expansion and intensification of agricultural activity has been very marked, causing accelerated fragmentation of the landscape [10] and consequent losses in biodiversity.
There are contributions to the arthropod communities of agroecosystems in different provinces of our country; for example, a study carried out in Entre Ríos estimates the biodiversity of arthropods to implement conservation strategies [11] in the same province studies of diversity and abundance were carried out in agroecosystems [10, 12] and a particular soybean crop [11]. In the province of Santa Fe, diversity was also analyzed in different crops, such as soybeans [13].
This research presents the taxonomic richness of the understory entomofauna present in the FCV-UNR property, understood as a complex (vertical variation) and heterogeneous (horizontal variation) environment in terms of plant diversity, to be used as a baseline for future comparative work with environments that have experienced intense transformations of the original vegetation cover, becoming more homogeneous environments with a simplified plant structure (agroecosystems).
MATERIALS AND METHODS
Area of study
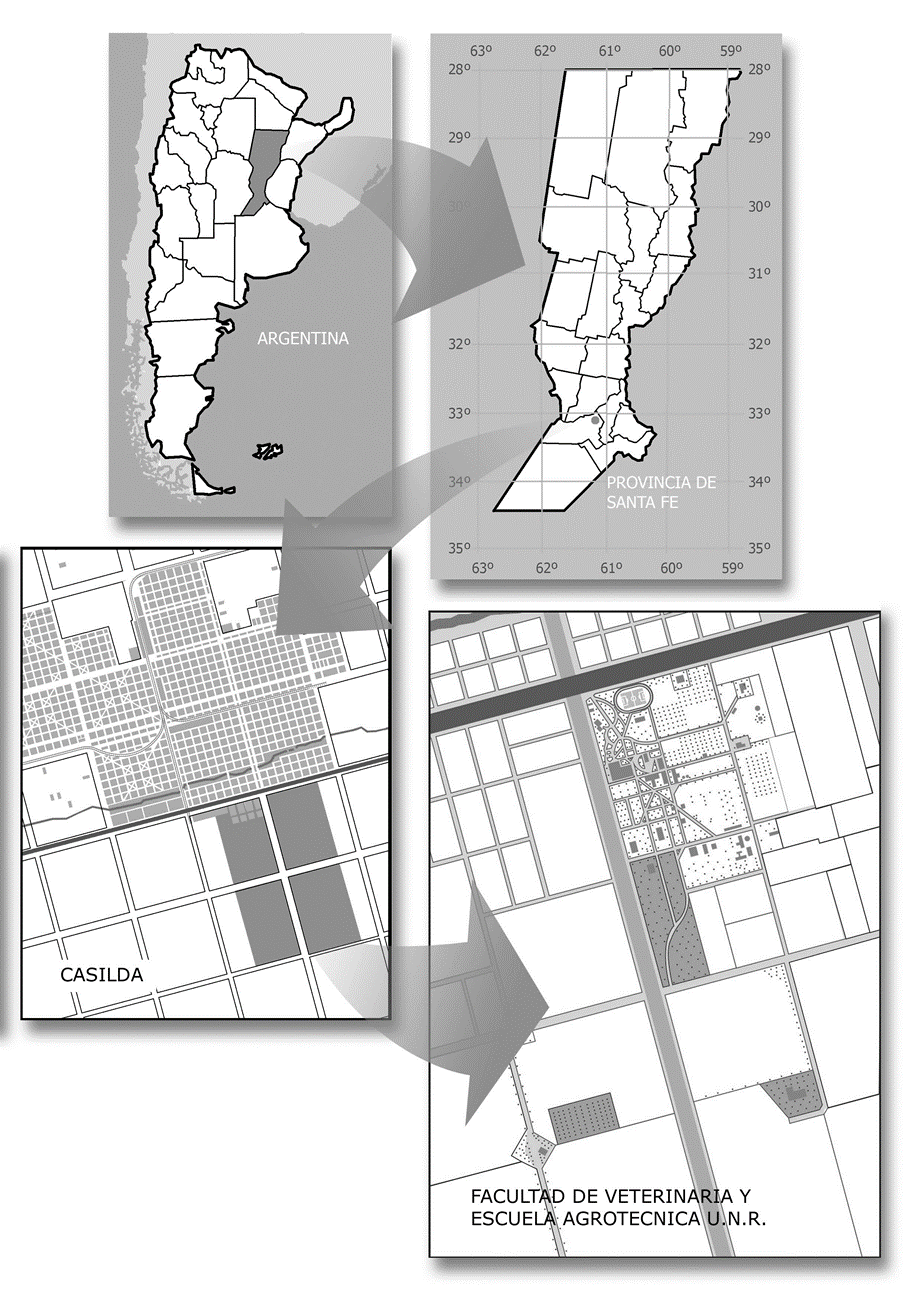
The Faculty of Veterinary Sciences of the National University of Rosario is located in Casilda, head of the Caseros Department, in the south of the province of Santa Fe. Its property covers an area of approximately 240 hectares and was declared a "Natural Protected Area" in 2007 (CD Resolution Nº 188/07) due to its leading role as a refuge for wildlife in a purely agricultural area (Figure 1).
The climate in this region is temperate, with average temperatures between 14 and 20 °C [14]. Precipitation varies throughout the year, being more intense in spring and summer. Before the region was almost entirely dominated by livestock and agricultural production, dense and extensive grasslands predominated [15].
|
|
|
Figure 1. Faculty of Veterinary Sciences - National University of Rosario, Casilda, Santa Fe, Argentina. |
The prescribed sampling was carried out between June (autumn-winter) and December (spring-summer) of 2022. Given the heterogeneity of the study forest, markedly different microsites were selected as sampling areas. Two homogeneous sampling areas were selected in each microsite to capture the entomofauna under study.
The microsites were established as follows:
|
|
|
a) |
|
|
|
b) |
|
|
|
c) |
|
|
|
d) |
|
|
|
e) |
|
Figure 2. Microsites that made up the study area on the premises of the Faculty of Veterinary Sciences of the National University of Rosario. |
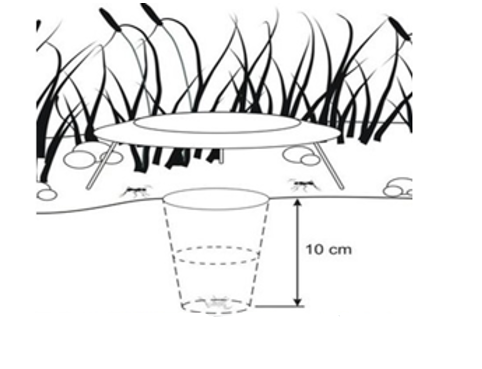
Two plastic pitfall traps (diameter, 8 cm; depth, 10 cm) were placed in each sampling area (Figure 3). The traps were filled with 200 mL of diluted ethylene glycol (20%) and a drop of detergent to reduce surface tension. Ethylene glycol prevents evaporation and preserves the captured organisms. The traps remained active in the field for 5 consecutive days for each month of sampling, accounting for a total trapping effort of 300 trap days [5 areas/microsite × 2 trap/area × 6 months × 5 days/month = 300 trap days].
The collected material was preserved in 70% alcohol for later determination.
|
|
|
a) |
|
|
|
b) |
|
|
|
c) |
|
|
|
d) |
|
|
|
e) |
|
Figure 3. a, b, c) Fieldwork, d) laboratory, and e) pitfall trap scheme. |
Total diversity (gamma diversity) was estimated following [16], who define it as the number of morphospecies in the set of sites that make up the study area; in this case, the number of morphospecies recorded in the different microsites present in the study area.
In addition, the following were determined: (a) taxonomic richness (S), understood as the number of morphospecies in a sample; (b) relative abundance, estimated as the percentage fraction of the total number of morphospecies; (c) alpha diversity (intra-area/microsite), considering specific richness and structure. The latter was determined according to Shannon and Wiener's diversity index, which quantifies the total diversity of a sample, being influenced by two fundamental components: richness and equity. It thus considers the importance value of each morphospecies and expresses the uniformity of importance values across all species in the sample. The formula for this function is H'= -Σ (pi x log2 pi), where pi is the proportion of the total number of individuals in the sample that corresponds to the species, whose values are displayed between zero when there are only one species, and the maximum (H'max) corresponding to log2 S.
The degree of similarity in morphospecies composition among the different areas/microsites was estimated using Jaccard's index [17].
The names of the higher taxa were given according to Borror et al. [18], while for most genera and species, the classification used by Morrone and Coscarón [19] and Claps et al. [20] was followed. The collected material was identified to the order and family level; even the species category was determined when possible. The rest of the material was identified as recognizable taxonomic units or "morphospecies" different from the above. Determining the species level is time-consuming and sometimes practically impossible due to the lack of expert taxonomists for the groups involved. Taxonomic detail often does not improve the ecological results obtained despite the time required to acquire this knowledge [21].
The trophic structure of the community was studied by assigning each morphospecies captured to a trophic group according to the information available in the literature. Four major groups are defined: herbivores, predators, detritivores, and ants. Ants are considered a separate group because most species exploit diverse food sources opportunistically [22].
RESULTS AND DISCUSSION
A total of 2631 individuals were captured and grouped into five classes, 15 orders, and 43 families (Figure 4, Table 1). Of the 68 morphospecies identified, 51.47% (35 sp.) were determined at the specific level, 22.05% (15 sp.) at the generic level, 25% (17 sp.) at the family level, and 1 sp. as morphospecies different from the above. Based on these results, the gamma diversity for the understory of the Faculty of Veterinary Sciences of the National University of Rosario was established at 68 morphospecies.
Table 1. The taxonomic richness of the entomofauna present in the microsites surveyed on the premises of the Faculty of Veterinary Sciences of the National University of Rosario.
|
FAMILIA |
Especie/ Morfoespecie |
GT |
Sampling sites/microsites |
|||||||||
|
Autumn-winter |
Spring-festival |
|||||||||||
|
1 |
2 |
3 |
4 |
5 |
1 |
2 |
3 |
4 |
5 |
|||
|
ARANEIDAE |
Araneidae sp. |
PRE |
4 |
1 |
2 |
4 |
2 |
2 |
9 |
11 |
||
|
Alpaida gallardoi |
PRE |
6 |
||||||||||
|
CLUBIONIDAE |
Clubionidae sp. |
PRE |
3 |
3 |
||||||||
|
GNAPHOSIDAE |
Gnaphosidae sp. |
PRE |
2 |
3 |
||||||||
|
LYCOSIDAE |
Lycosidae sp. |
PRE |
3 |
1 |
7 |
3 |
4 |
9 |
16 |
|||
|
TETRANYCHIDAE |
Tetranychidae sp. |
PRE |
1 |
1 |
||||||||
|
SCLEROSOMATIDAE |
Holmbergiana weyenberghi |
PRE |
3 |
5 |
||||||||
|
GONYLEPTIDAE |
Pachyloides thorellii |
PRE |
2 |
|||||||||
|
SCOLOPENDRIDAE |
Rhysida sp. |
PRE |
1 |
2 |
||||||||
|
PSEUDONANNOLENIDAE |
Pseudonannolene meridionalis |
DET |
1 |
1 |
8 |
5 |
1 |
12 |
10 |
|||
|
ARMADILLIDAE |
Armadillidium vulgare |
DET |
216 |
364 |
631 |
166 |
147 |
|||||
|
PORCELLIONIDAE |
Porcellio laevis |
DET |
1 |
11 |
5 |
|||||||
|
BLATIDAE |
Blattidae sp. |
DET |
22 |
19 |
12 |
5 |
26 |
|||||
|
ENTHOMOBRYIDAE |
Enthomobryidae sp. |
DET |
28 |
14 |
16 |
54 |
76 |
|||||
|
CARABIDAE |
Calosoma argentinense |
PRE |
3 |
1 |
1 |
|||||||
|
Calosoma granulatum |
PRE |
1 |
||||||||||
|
Galerita collaris |
PRE |
8 |
4 |
|||||||||
|
Clivina platensis |
PRE |
1 |
||||||||||
|
Blennidus loxandroides |
PRE |
6 |
8 |
3 |
||||||||
|
Argutoridius bonariensis |
PRE |
2 |
12 |
6 |
1 |
|||||||
|
Trirammatus striatulus |
PRE |
1 |
11 |
4 |
7 |
|||||||
|
Pterostichini sp. |
PRE |
2 |
2 |
1 |
||||||||
|
Arthrostictus chlaenioides |
PRE |
1 |
||||||||||
|
Notiobia cupripennis |
PRE |
2 |
1 |
|||||||||
|
Carabidae sp. (larvas) |
PRE |
3 |
1 |
1 |
||||||||
|
CURCULIONIDAE |
Listroderes apicalis |
HER |
4 |
2 |
1 |
|||||||
|
ELATERIDAE |
Conoderus bellus |
HER |
3 |
1 |
||||||||
|
Heteroderes laurentii |
HER |
8 |
11 |
|||||||||
|
Elateridae sp. (larvas) |
HER |
1 |
1 |
1 |
||||||||
|
NITIDULIDAE |
Nitidulidae (1 sp.) |
DET |
16 |
8 |
11 |
12 |
4 |
|||||
|
SCARABAEIDAE |
Aphodius sp. |
HER |
6 |
4 |
7 |
1 |
2 |
|||||
|
STAPHYLINIDAE |
Staphylinidae sp. |
PRE |
8 |
9 |
4 |
1 |
||||||
|
FORFICULIDAE |
Doru sp. |
PRE |
1 |
|||||||||
|
ANTHOMYIIDAE |
Anthomyia punctipennis |
DET |
1 |
1 |
6 |
3 |
||||||
|
BIBIONIDAE |
Dilophus sp. |
HER |
1 |
2 |
||||||||
|
CECIDOMYIIDAE |
Cecidomyiidae sp. |
HER |
14 |
2 |
17 |
2 |
||||||
|
CHIRONOMIDAE |
Chironomidae sp. |
DET |
8 |
5 |
3 |
|||||||
|
DROSOPHILIDAE |
Drosophila melanogaster |
DET |
1 |
3 |
2 |
|||||||
|
LIMONIIDAE |
Limoniidae sp. |
? |
5 |
3 |
||||||||
|
PHORIDAE |
Phoridae sp. |
DET |
32 |
4 |
20 |
7 |
16 |
|||||
|
MUSCIDAE |
Bithoracochaeta calopus |
DET |
5 |
3 |
1 |
2 |
1 |
8 |
1 |
4 |
||
|
Limnophora sp. |
DET |
1 |
2 |
2 |
11 |
|||||||
|
Musca domestica |
DET |
1 |
9 |
3 |
1 |
|||||||
|
Syllimnophora sp. |
DET |
5 |
9 |
2 |
||||||||
|
Psilochaeta chalybea |
DET |
5 |
||||||||||
|
STRATIOMYIDAE |
Hermetia sp. |
DET |
2 |
|||||||||
|
SYRPHIDAE |
Allograpta sp. |
PRE |
1 |
2 |
4 |
|||||||
|
S/D |
Diptera sp. |
DET |
2 |
1 |
3 |
1 |
3 |
|||||
|
FORMICIDAE |
Linepithema humile |
HORM |
6 |
4 |
1 |
13 |
11 |
6 |
9 |
19 |
||
|
Acromyrmex lundi |
HORM |
2 |
7 |
1 |
8 |
3 |
9 |
|||||
|
Camponotus mus |
HORM |
1 |
1 |
6 |
||||||||
|
Hypoponera argentina |
HORM |
3 |
2 |
4 |
||||||||
|
Dorymyrmex brunneus |
HORM |
5 |
||||||||||
|
Pseudomyrmex gracilis |
HORM |
1 |
1 |
3 |
||||||||
|
ICHNEUMONIDAE |
Pimpla sp. |
PRE |
1 |
2 |
||||||||
|
SCOLIIDAE |
Campsomeris sp. |
PRE |
2 |
|||||||||
|
VESPIDAE |
Polistes sp. |
PRE |
2 |
2 |
3 |
|||||||
|
Isodontia sp. |
PRE |
1 |
1 |
|||||||||
|
APIDAE |
Apis mellifera |
HER |
1 |
4 |
||||||||
|
PENTATOMIDAE |
Nezara viridula |
HER |
3 |
1 |
2 |
|||||||
|
CICADIDAE |
Cicadidae sp. |
HER |
1 |
|||||||||
|
LYGAEIDAE |
Lygaeus alboornatus |
HER |
3 |
|||||||||
|
NABIDAE |
Nabidae sp. |
PRE |
1 |
1 |
||||||||
|
BERYTIDAE |
Jalysus sp. |
HER |
1 |
3 |
||||||||
|
NOCTUIDAE |
Spodoptera frugiperda |
HER |
1 |
|||||||||
|
Agrotis malefida |
HER |
2 |
||||||||||
|
GRYLLIDAE |
Acheta assimilis |
HER |
2 |
1 |
1 |
8 |
1 |
1 |
5 |
9 |
||
|
GRYLLOTALPIDAE |
Neoscapteriscus sp. |
HER |
2 |
2 |
||||||||
References: GT (trophic group), HER (herbivores), PRE (predators), DET (detritivores), and HORM (ants). Ants are considered a separate group because most species exploit diverse food sources opportunistically [22].
The highest richness corresponded to the Class Insecta, with 56 (82.35%) morphospecies of 9 orders and 33 families. The abundance of this class represented 35.80% of the total number of individuals collected, with the morphospecies Enthomobryidae sp. contributing the most individuals, 19.95% (n = 188). The Class Malacostraca, with only two species, presented 58.57% of the total abundance, with a marked dominance of Armadillidium vulgare (n = 1524, 98.89%). The eight morphospecies of the Class Arachnida reached 4.06% of the total abundance, with a dominance of Lycosidae sp. The Classes Diplopoda and Chylopoda abundance was 1.44 and 0.11%, respectively.
|
|
|
a) |
|
|
|
b) |
|
|
|
c) |
|
|
|
d) |
|
|
|
e) |
|
|
|
f) |
|
Figure 4. a) Armadillidium vulgare, b) Enthomobryidae sp., c) Linepithema humile, d) Lycosidae sp., e) Cecidomyiidae sp., and f) Phoridae sp. |
Concerning diversity by a microsite, which was determined from the Shannon-Wiener index, microsites 5 and 1 were found to be the most biodiverse with H' = 2.51 and 2.48, respectively. Microsite 4 with H' = 2.13 is very close in terms of diversity. The microsites with the lowest diversity were 2 with H' = 1.67 and 3 with H' = 1.36.
Concerning the results obtained from the Jaccard index, it is important to mention that similarity is based on the incidence of species in each microsite (presence or absence). Hence, the results can be visualized as the proportion or percentage of shared morphospecies, allowing us to know in detail the similarity between previously defined communities.
Taking into account that a value closer to 1 indicates greater similarity [23], it can be said that there was low similarity between all the microsites studied (Table 2). In all cases, values were far from 1 (0.28-0.59), the most related areas being microsite 2 and microsite 3 (Jaccard index = 0.59) with 28 shared morphospecies; the other pairs showed lower similarity values. These low similarities indicate that most species from one community do not occur in the other.
Table 2. Jaccard index for biota pairs present in the microsites surveyed at the Faculty of Veterinary Sciences, National University of Rosario.
|
MICROSITES |
Microsites 1 |
Microsites 2 |
Microsites 3 |
Microsites 4 |
Microsites 5 |
|
Microsites 1 |
1 |
0.53 |
0.58 |
0.4 |
0.4 |
|
Microsites 2 |
0.53 |
1 |
0.59 |
0.35 |
0.28 |
|
Microsites 3 |
0.58 |
0.59 |
1 |
0.33 |
0.31 |
|
Microsites 4 |
0.4 |
0.35 |
0.33 |
1 |
0.52 |
|
Microsites 5 |
0.4 |
0.28 |
0.31 |
0.52 |
1 |
Pitfall traps represent the most commonly employed sampling method for collecting arthropods that move along the soil surface [24]. However, captures obtained by this method have been found to represent surface activity more than specific population levels of the various species [25].
In this study, the number and diameter of the traps used are similar to those used in previous research conducted in similar environments [25-27]. It is estimated that these conditions allow a reasonable approximation of the richness and abundance of the surface epigean fauna in the different study environments.
The number of insects captured by pitfall traps directly correlates with the perimeter of the traps. It has been observed that, at a constant trap density, an increase in trap diameter translates into an increase in both abundance and diversity of Carabids and spiders [28], as well as ants [29]. Work et al. [28] suggest that, to ensure the capture of larger species, especially those larger than 10 mm, it is advisable to use traps with a diameter greater than 10 cm.
This sampling method has the disadvantage of depending on the population density and the activity rate of the various species. The locomotor activity of the organisms is subject to several factors, such as weather conditions or the physical characteristics of the surrounding terrain [30]. Generally speaking, increased mobility is observed with increasing mean temperature, while activity tends to decrease in rainy conditions. Disparities in terrain roughness, caused by the accumulation of dry matter on the surface or variations in vegetation architecture, may influence capture rates in pitfall traps.
Despite these challenges, pitfall traps remain a fast, effective, and economical sampling method for conducting biodiversity surveys in various environments.
Classifying samples as recognizable taxonomic units or morphospecies is generally regarded as a reliable approach for conducting ecological biodiversity or conservation biology studies [31]. It saves time and circumvents serious drawbacks produced by the lack of expert taxonomists who specialize in the various groups of arthropods captured. However, in many cases, there are overestimates in the number of species involved, so errors in classification can be high and cause severe problems in the analysis of the data obtained [32].
A significant effort was made to achieve taxonomic determination to the specific level of a high percentage of the captured material to address these limitations. Some of the material was sent to specialists for identification; however, the large number of species and taxa involved partially made the task difficult.
The richness, abundance, and specific composition of arthropods are intrinsically linked to the structure of the landscape. The spatial distribution of these organisms is strongly influenced by the configuration of the surrounding environment, and it is evident that in areas where the landscape has been significantly simplified due to anthropogenic activities, such as intensive agriculture, there are substantial modifications in the abundance and diversity of the associated arthropod communities [33]. Different species exhibit diverse responses to environmental disturbances, leading to both increased abundance and local rarefaction or extinction in affected environments [34].
There is a consensus that less disturbed environments, such as vegetation patches, natural corridors, habitat edges, and uncultivated plots, play a key role not only in increasing landscape heterogeneity but also in the ability to maintain and increase biological diversity in agroecosystem-dominated landscapes [26]. These less disturbed microhabitats provide refuge and resources for diverse species, thus contributing to sustainability and biodiversity in areas dominated by intensive agricultural practices.
The understory that characterizes the grounds of the Faculty of Veterinary Sciences of the National University of Rosario is an exceptionally stable environment throughout the year. The surface layer of leaf litter and the shade generated by the vegetation play a crucial role by acting as a buffer against extreme weather conditions. This dynamic stabilizes by retaining moisture during dry winters and reducing surface temperature in summer, creating microclimates conducive to soil life [35]. Such microenvironments provide safe refuges for feeding reproduction and are used by epigean species significantly.
This stable and relatively undisturbed context provides a favorable habitat for the persistence and development of soil fauna, highlighting the importance of conserving areas with these characteristics to maintain biodiversity and ecosystem functionality.
The above mentioned is consistent with the results obtained in this research, where beyond the diversity obtained, which was unknown until now for this study area, the two most abundant morphospecies, Armadillidium vulgare and Enthomobrydae sp. a morphospecies of the Order Collembola, which represents 65.07% of the total number of individuals collected, provide important ecosystem services such as decomposition of organic matter, improvement of soil structure, pest control, nutrient cycling, and soil aeration, among others.
Springtails are panphytophagous and can feed on decaying organic matter, so they are considered detritivores, although depending on conditions, their food preferences may vary. However, most ingest pollen, algae, spores, and fungal mycelia [36]. Within the group are taxa susceptible to environmental variations, i.e., they have only been found in ecological situations that did not vary abruptly. This indicator characteristic of Collembola is one of the reasons why they have been studied in situations subjected to anthropogenic impact [37].
For its part, Armadillidium vulgare (Latreille, 1804; Isopoda: Oniscidea) is considered a potential bioindicator of soil and agroecosystem quality due to its wide geographical distribution (cosmopolitan), easy taxonomic identification and for being the dominant component in the guild of detritivores in temperate regions [38].
A. vulgare populations are very sensitive to pesticide application and tillage practices, showing differences in density between conventional and organic crops and differences in biomass, being higher under no-till or minimum tillage conditions [39]. Insecticides and herbicides increase mortality rates and decrease growth and fecundity rates by reducing the nutritional quality of leaf litter [40]. Mortality also results from habitat simplification (structure) and reduced shelter capacity, as with many tillage practices [41].
CONCLUSION
This exploratory, descriptive research has provided an important baseline on the epigean entomofauna inhabiting the understory of the FCV-UNR property. An exhaustive fieldwork and collection of 2631 individuals has identified a taxonomic richness of 68 morphospecies. The diversity index obtained, H' = 2.12, provides a quantitative measure of the varied spectrum of species present in this particular ecosystem and serves as an input to assess the ecosystem's overall health [42]. This indicator further reveals the complexity and interconnectedness of the different life forms coexisting in the understory, serving as a key indicator for assessing ecological stability and resilience [43].
The role of soil organisms has received little attention because their services as ecosystem engineers, litter fragments, and regulators of biotic stress have been replaced by operations and techniques dependent on non-renewable energy sources [44]. However, the diversity, number, and functions of soil invertebrates are sensitive to stress and environmental change associated with tillage, fertilizer and pesticide application, burning, logging, and other cultivation practices that impact soil faunal communities by altering organic matter inputs and modifying microhabitats in terms of the physical and chemical quality of soils [45, 46]. The magnitude of the effects of land use on edaphic fauna depends on the type of use, the sowing system (conventional or direct), crop diversity and rotation, and the inputs used [47].
These results are essential for understanding local biodiversity and lay the groundwork for future comparative studies. The possibility of contrasting these findings with highly anthropized or simplified ecosystems, such as the agroecosystems that constitute the dominant matrix in the current landscape of southern Santa Fe, offers a unique opportunity to evaluate the impact of human activity on regional biodiversity. The information gathered in this research is a valuable tool to guide the region's conservation and sustainable management strategies, providing key perspectives for environmental management and biodiversity preservation [48].
Finally, it is imperative to understand the importance of considering biological diversity in the planning and implementing conservation measures, with direct implications for decision-making in environmental politics at the local and regional levels.
ACKNOWLEDGMENTS: None
CONFLICT OF INTEREST: None
FINANCIAL SUPPORT: None
ETHICS STATEMENT: None