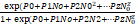Entomology and
Applied Science Letters
Applied Science Letters
 2024
Volume 11
Issue 2
2024
Volume 11
Issue 2

Phenacoccus solenopsis Tinsley is a polyphagous insect that has become a drastic threat to agriculture in subtropical and tropical regions of the world since the first decade of the 20th century. Aenasius bambawalei Hayat is an endoparasitoid that parasitizes the third instar nymphs of P. solenopsis. In the current research, the functional response of this parasitoid to sublethal doses of spirotetramat and chlorpyriphos was investigated on third-instar nymphs of P. solenopsis at a 27 ± 1 °C temperature, relative humidity of 65 ± 5%, and a 10:14 h light-dark cycle. The findings of logistic regression revealed that the functional response in all three treatments of control, chlorpyriphos, and spirotetramat was of the second type. The search rate (a) of the bee in the control, spirotetramat, and chlorpyriphos treatments was calculated as 0.2609, 0.1107, and 0.0556 times per hour, respectively, which was 57% and 78% lower than the control in the spirotetramat and chlorpyriphos treatments, respectively. The access time (Th) in the control, spirotetramat, and chlorpyriphos treatments was calculated as 5.018, 6.335, and 9.390 hours, respectively. In the spirotetramat and chlorpyriphos treatments, the access time was about 26% and 87% higher than the control treatment, respectively. The access rate was also determined as 4.63, 3.28, and 2.55 host nymphs in the control, spirotetramat, and chlorpyriphos treatments, respectively, which was 31% and 45% lower than the control in the spirotetramat and chlorpyriphos treatments. The findings of this research revealed that the insecticides spirotetramat and chlorpyriphos, even at low doses, hurt the functional response behavior of bees and can cause disruption in biological control.
INTRODUCTION
Phenacoccus solenopsis Tinsley is a major pest of cotton growing areas in tropical and subtropical regions of the world and has caused significant economic losses to cotton crops in Pakistan and India [1-3]. This weevil, by forming colonies on the plant and sucking plant sap, reduces crop quality and causes plant growth arrest, leaf and fruit deformation and shedding, the transmission of viral diseases and ultimately plant death. The pest also attracts sooty molds by producing large amounts of honeydew and affects photosynthesis, respiration, and other biological activities of the plant [4]. In recent years, this pest has been reported from various other hosts in addition to cotton and has been collected from more than 200 plant host species so far [5-7].
Chemical control of this pest is not cost-effective, either economically or in terms of spreading chemicals and eliminating natural enemies [8, 9]. Also, chemical control of this pest, like other thrips, is not successful due to the presence of a waxy coating on the body that disrupts the effectiveness of chemical pesticides [10]. Continuous use of chemical pesticides will cause pest reversibility, resistance in insects, environmental pollution, and harmful effects on human health [11]. In controlling this pest, their natural enemies (parasitoids and predators) are of particular importance and a large number of them have been identified [8, 9].
Parasitoids are the most successful and important group of natural enemies utilized in the biological control of insects. The parasitoid wasp Aenasius bambawalei Hayat from the order Hymenoptera and the family Encyrtidae is the most important internal parasite of P. solenopsis. This wasp is a solitary parasitoid of mealybug nymphs and in some areas such as northern India, it is the only dominant and invasive parasitoid that successfully controls this weevil [11]. This wasp was first reported from India in 2009 and is responsible for the reduction of mealybug populations in northern India [12, 13]. This parasitoid wasp has been reported from India, Pakistan [14], China [15], Australia [16], and Iran [17].
In the world, several insecticides are used to control P. solenopsis on various crops. Although these insecticides are used to control the weevil, they are likely to be harmful to the weevil’s natural enemies [18]. Information on the impact of sublethal doses of insecticides on the biology and behavior of the natural enemies of P. solenopsis is limited. The relative toxicity of 10 common insecticides on P. solenopsis and the parasitoid A. bambawalei was investigated by Nagrare et al. [11]. Another study investigated the toxicity of the insecticides endosulfan, monocrotophos, profenofos and dimethoate to the parasitoid wasps Aenasius advena Compere and A. bambawalei [18]. The effect of the insecticides cypermethrin, azadirachtin, and pyrazinyl on the functional response of the parasitoid wasp Habrobracon hebetor (Hym.: Braconidae) was investigated [19]. The effect of the two toxins imidacloprid and buprofezin on the functional response of Eretmocerus mundus Mercet, a parasitoid wasp of cotton white boll nymphs, has been investigated [20]. Rostami et al. [21] also studied the functional response of the parasitoid wasp H. hebetor larvae under the influence of the toxins azadirachtin and flubendiamide.
A review of the available literature showed that despite the use of various chemical toxins to control P. solenopsis, no study has been conducted so far on the impact of sublethal concentrations of common chemical toxins on the behavior of the parasitoid wasp A. bambawalei. Therefore, this study proposes to study the functional response of this wasp under the influence of sublethal concentrations of spirotetramat and chlorpyriphos.
MATERIALS AND METHODS
Insect rearing
P. solenopsis mealybugs were obtained from infested Chinese marshmallow bushes. Nymphs and adults were transferred from the infested leaves to potato sprouts in a ventilated container measuring 8 × 14 × 20 cm with a brush, thus forming a colony of mealybugs. The containers containing mealybugs were placed in an incubator at a 25 ± 2 °C temperature and a 65 ± 5% relative humidity for rearing. Leaves containing mummified mealybugs were collected from Chinese marshmallow bushes, and parasitized mealybugs were separated with a brush and placed in ventilated containers. After the emergence of adult parasitoid wasps and their identification, they were transferred to containers containing mealybug-infested potatoes. Small drops of a solution of water and honey were placed on the walls of the containers daily to feed the bees.
Insecticides used in the experiments
In this study, the insecticides spirotetramat suspension 10% and chlorpyriphos suspension 40.8% were used. After conducting preliminary tests and based on a 10% field dose of each pesticide, concentrations of 1 ppm chlorpyriphos and 7.5 ppm spirotetramat were considered for the functional response of the parasitoid wasp.
Investigation of the sublethal impacts of pesticides on the functional response of parasitoid wasps
To investigate the effect of 1 ppm and 7.5 ppm concentrations of chlorpyriphos and spirotetramat on the functional response of the parasitoid wasp, properly ventilated cylindrical plastic containers with a height of 15 and a diameter of 7 cm were impregnated with the pesticides. Distilled water was used in the control treatment. The treatments were exposed to air for 24 hours. Then, 40 to 50 pairs of one-day-old parasitoid wasps were released into the containers impregnated with the pesticides. After 24 hours, several surviving females and males were randomly selected and each pair was released into containers containing potato sprouts with specific densities (2, 4, 8, 16, 32, and 64) of third-instar nymphs of the weevil. Bees were kept on the thrips for 24 hours and then removed from the test containers. After the bee pupae were identified, the parasitized nymphs number in each treatment was counted. Parasitization of nymphs was determined based on the formation of mummies. In this experiment, 10 replicates were considered for each insecticide and control treatment.
Functional response data analysis
To determine the type of functional response and its parameters, SAS software and the two-stage method proposed by Juliano (2001) were used.
The first stage included determining the type of functional response using logistic regression of the ratio of parasitized hosts (Ne) to the number of initial hosts (N0) through a polynomial function.
|
|
(1) |
P0, P1, P2, and Pz are parameters determined by the CATMOD method. The sign of the coefficient parameter N0 (linear part of the curve) in the regression analysis table determines the functional response type. The negative sign of the linear part indicates that the curve is initially decreasing and the functional response is of the second type, and the positive sign shows that the curve is initially increasing and the functional response is of the third type.
The second step is to estimate the functional response parameters and compare them in different treatments. After determining the type of functional response and selecting the appropriate model, the nonlinear least squares regression of the number of parasitized hosts (Ne) to the number of initial hosts (N0) was used to estimate the parameters (NLIN method in the SAS statistical program). After estimating the search power (a) and access time (Th) parameters, these parameters were compared in different treatments and used as a criterion for evaluating the efficiency of the parasitoid. The search power and access time parameters were evaluated by comparing the upper and lower limits in different treatments. According to the type of functional response in this experiment, the following relationship was used to estimate the search power and access time parameters:
|
Ne = N0 {1-exp[a(ThNe − T)]} |
(2) |
Parameter a or search power is the ratio of hosts that the parasitoid encounters per unit of search time and Th or access time includes all operations other than search, including chasing and surrendering, eating prey or tentacles on the host, parasitizing, cleaning, self- The resting period is the period between the predator's feeding or the parasitoid's spawning, which includes the time between observing a prey or host and moving towards it to feed or parasitize until the search for a new prey or host is resumed. The maximum attack rate in predators, or the maximum number of hosts that can be attacked, is calculated from the relationship T/Th. In addition to Th, the parasitoid egg limit also determines the parasitism rate.
RESULTS AND DISCUSSION
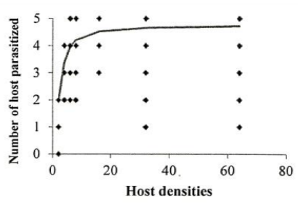
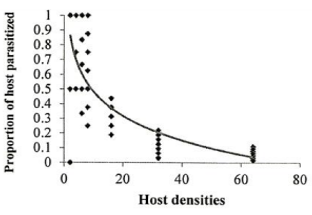
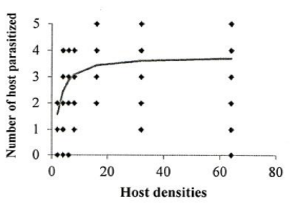
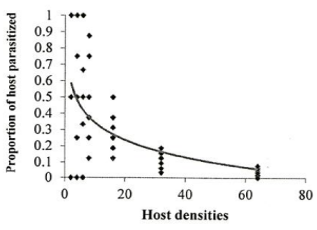
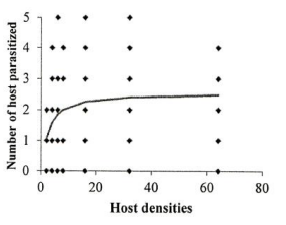
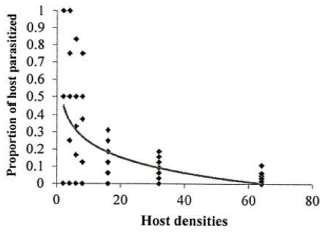
Investigation of the sublethal impacts of chlorpyriphos and spirotetramat pesticides on the functional response of the parasitoid bee A. bambawalei. It was shown that the slope of the line portion of the curve in all three treatments of control, spirotetramat, and chlorpyriphos was negative and was calculated as -0.2123, -0.0328, and -0.2055, respectively. Therefore, the functional response of the bee in all three treatments tested was of the second type (Table 1). In this type of functional response, with increasing thrips density (2, 4, 6, 8, 16, and 32), the number of parasitized hosts enhanced, but the ratio of parasitized nymphs to the initial host density reduced with increasing host density and finally, the curve became asymptotic (Figure 1).
Table 1. Findings from fitting logistic regression of third-instar nymph parasitized proportion by the A. bambawalei against the initial number of nymphs in the treatments of chlorpyriphos and spirotetramat pesticides and the control treatment.
|
Treatments |
Parameters |
Estimate |
SE |
P |
|
Control |
Constant (P0) |
1.8229 |
0.3823 |
< 0.0001 |
|
Linear (P1) |
– 0.2123 |
0.0616 |
0.0006 |
|
|
Quadratic (P2) |
0.00375 |
0.00240 |
0.1186 |
|
|
Cubic (P3) |
– 0.00002 |
0.000024 |
0.3219 |
|
|
Spirotetramat |
Constant (P0) |
0.1096 |
0.3440 |
0.7501 |
|
Linear (P1) |
– 0.0328 |
0.05840 |
0.5745 |
|
|
Quadratic (P2) |
0.00217 |
0.00234 |
0.3551 |
|
|
Cubic (P3) |
– 0.00030 |
0.000024 |
0.2142 |
|
|
Chlorpyriphos |
Constant (P0) |
0.2930 |
0.3843 |
0.4458 |
|
Linear (P1) |
– 0.2055 |
0.0692 |
0.0030 |
|
|
Quadratic (P2) |
0.00527 |
0.00278 |
0.0581 |
|
|
Cubic (P3) |
– 0.00006 |
0.000028 |
0.1034 |
|
|
|
|
a) |
|
|
|
|
|
b) |
|
|
|
|
|
c) |
|
|
Figure 1. Functional response of A. bambawalei females to various densities of third-instar nymphs of P. solenopsis in the control, a) spirotetramat, b) and chlorpyriphos, c) treatments (figures on the left show the number of parasitized hosts and figures on the right show the proportion of parasitized hosts). |
|
Table 2. Parameters of the second type of functional response of A. bambawalei females to third-instar nymphs of P. solenopsis in the three treatments of control, chlorpyriphos, and spirotetramat.
|
Treatments |
Parameters |
Estimate |
Asymptotic SE |
Asymptotic 95% CI |
T/Th |
R2 |
|
|
Lower |
Upper |
||||||
|
Control |
a (h-1) |
0.2609 |
0.1578 |
–0.0540 |
0.5759 |
4.63 |
0.88 |
|
Th (h) |
5.0187 |
0.3507 |
4.3189 |
5.7184 |
|||
|
Spirotetramat |
a (h-1) |
0.1107 |
0.0613 |
–0.0117 |
0.2331 |
3.28 |
0.77 |
|
Th (h) |
6.3353 |
0.6506 |
5.0371 |
7.6336 |
|||
|
Chlorpyriphos |
a (h-1) |
0.0556 |
0.0376 |
–0.0194 |
0.1306 |
2.55 |
0.61 |
|
Th (h) |
9.3903 |
0.4465 |
6.5036 |
12.2769 |
|||
The functional response parameters of A. bambawalei in three treatments of control, spirotetramat, and chlorpyriphos are shown in Table 2. The search rate (a) of bee in control, spirotetramat, and chlorpyriphos treatments was determined as 0.2609, 0.1107, and 0.0556 per hour, respectively, which was the highest in control and the lowest in chlorpyriphos treatment. The search rate in spirotetramat and chlorpyriphos treatments was 57% and 78% lower than the control, respectively. The access time (Th) in control, spirotetramat and chlorpyriphos treatments was calculated as 5.018, 6.335, and 9.390 hours, respectively, which was about 26% and 87% higher than the control in spirotetramat and chlorpyriphos treatments, respectively. The acquisition rate was also determined to be 4.63, 3.28, and 2.55 host nymphs in the control, spirotetramat, and chlorpyriphos treatments, respectively, which was 31% and 45% lower than the control in the spirotetramat and chlorpyriphos treatments.
The biological control method's success is associated with the choice of an appropriate biological control agent with satisfactory control of the target pest and its compatibility with other control methods. Burks et al. [22]. Control of P. solenopsis is mainly associated with the use of insecticides on the foliage. Therefore, there is a possibility that natural enemies will be exposed to pesticide residues when searching on the leaves of sprayed plants. According to the findings of the present research, the functional response of the parasitoid wasp A. bambawalei in the control treatment and the insecticides spirotetramat and chlorpyriphos were of the second type. The second type of functional response for this parasitoid was observed in the studies of Feng et al. [23]. This result indicated that insecticides did not affect the type of parasitoid functional response. The type of functional response of natural enemies is affected by biotic and abiotic factors such as the parasitoid species studied, temperature, experimental conditions, host species, host plant, age of the parasitoid, and pesticides [24-26].
In studies conducted on the insecticide's impact on the functional response type of parasitoid wasps, similar results have been reported that insecticides do not impact the functional response type of parasitoids. In studies by Rashidi et al. [27], sublethal doses of the toxins phosalone, fipronil, pyriproxyfen, and diazinon did not impact the functional response type of the parasitoid wasp Habrobracon hebetor Say (Hymenoptera: Braconidae) compared to the control treatment. Also, sublethal concentrations (LC25) of the insecticide imidacloprid did not affect the functional response type of the parasitoid wasp Eretmocerus mundus Mercet (Hymenoptera: Aphelinidae) [20]. However, in the studies of the same researchers, the functional response type changed from type III to type II when parasitoids were affected by the recommended field dose of buprofezin. Faal–Mohammadali et al. [28] also reported in their study of the effect of chlorpyrifos and fenpropathrin insecticides on the functional response of the parasitoid wasp H. hebetor to various densities of Ephestia kehniella Zeller larvae that the type of parasitoid functional response was type III in the control and type II in the treatments, which indicated the effect of insecticides on changing the type of parasitoid functional response and was inconsistent with the findings of this research.
In this study, the residues of spirotetramat and chlorpyriphos insecticides affected the numerical value of the parameters of functional response of the bee A. bambawalei and caused an increase in the bee's access time and a reduction in the search rate. The impact of insecticides on the functional response parameters of other natural enemies has been reported in previous studies by researchers. In the studies of He et al. [29], a sublethal dose of imidacloprid caused a significant increase in the access time of Serangium japonicum Chapin (Coleoptera: Coccinellidae), a predator of cotton boll white eggs. In the studies of Rashidi et al. [27], parameters of the functional response of H. hebetor, including search power, were also affected by sublethal doses of insecticides phosalone, fipronil, pyriproxyfen, and diazinon. However, the negative impact of sublethal doses of insecticides on the functional response parameters of parasitoids is not true in all cases. For example, the LD20 concentration of chlorpyrifos caused a 5.1-fold increase in the search power of Leptopilina heterotoma Thomson (Hymenoptera: Eucoilidae), a parasitoid of Drosophila melanogaster larvae [30]. In addition, the phenomenon of hormoligosis, in which the performance of an individual increases after exposure to chemical stress, or other stresses, has been stated in a wide range of organisms, including insects [31].
Studies on the pesticide's effect on functional responses can help to improve the integrated pest management programs' success and the natural enemies release [19]. The findings of this research revealed that the insecticides spirotetramat and chlorpyriphos can affect the functional response of the parasitoid wasp A. bambawalei and, consequently, the biological control programs' success. These results showed that chlorpyriphos treatment caused the greatest increase in the time of arrival of A. bambawalei. Based on the findings of this research, the use of spirotetramat and chlorpyriphos pesticides during the activity of beneficial insects such as A. bambawalei is not recommended.
CONCLUSION
In this study, this parasitoid functional response to sublethal doses of spirotetramat and chlorpyriphos was investigated on third-instar nymphs of P. solenopsis at a 27 ± 1 °C temperature, 65 ± 5% relative humidity, and a 10:14 h light-dark cycle. The findings of logistic regression revealed that the functional response in all three treatments of control, chlorpyriphos, and spirotetramat was of the second type. The search rate (a) of the bee in the control, spirotetramat, and chlorpyriphos treatments was determined as 0.2609, 0.1107, and 0.0556 times per hour, respectively, which was 57% and 78% lower than the control in the spirotetramat and chlorpyriphos treatments, respectively. The access time (Th) in the control, spirotetramat, and chlorpyriphos treatments was calculated as 5.018, 6.335, and 9.390 hours, respectively. In the spirotetramat and chlorpyriphos treatments, the access time was about 26% and 87% higher than the control treatment, respectively. The access rate was also determined as 4.63, 3.28, and 2.55 host nymphs in the control, spirotetramat, and chlorpyriphos treatments, respectively, which was 31% and 45% lower than the control in the spirotetramat and chlorpyriphos treatments. The findings of this study revealed that the insecticides spirotetramat and chlorpyriphos, even at low doses, hurt the functional response behavior of bees and can cause disruption in biological control.
ACKNOWLEDGMENTS: None
CONFLICT OF INTEREST: None
FINANCIAL SUPPORT: None
ETHICS STATEMENT: None