Entomology and
Applied Science Letters
Applied Science Letters
 2023
Volume 10
Issue 3
2023
Volume 10
Issue 3

This study looked at the morphology of M. cernyi's male reproductive system to learn more about blister beetles in general. An aedeagus, two seminal vesicles, two vasa deferentia, an ejaculatory duct, three pairs of accessory glands, and two testes make up the male reproductive system of Mylabris cernyi. The morphology of this system was investigated using both optical and electron microscopy. There are 14–15 testicular follicles in the testis, arranged radially. In these testicular follicles, 3 different zones (growth, maturation, and differentiation) are distinguished from the periphery of the testis to its center. The testis emerges as a vas efferens later joining to form a vas deferens and seminal vesicle. Mature sperm masses are distinguished in their lumens. In M. cernyi, there are three pairs of accessory glands (Ag1, Ag2, and Ag3). Ag1 is short, tubular, and light yellow. It has columnar epithelial cells and a muscle layer. Its lumen contains basophilic secretory materials. Ag2 is long, coiled, tubular, and whitish, and its lumen has basophilic and fibrous secretory materials. A single layer of squamous epithelium envelops it. Ag3 is a long, tubular, coiled gland that secretes chemicals that are eosinophilic. The accessory gland opens into an ejaculatory duct. The ejaculatory duct wall is composed of a thick layer of muscle, epithelium, and intima. The intima layer's lumen-facing surface is covered in spines. The aedeagus is the opening of the ejaculatory duct. The findings offer the first morphological description of the male reproductive system of M. cernyi, which may be helpful for further investigation and investigations.
INTRODUCTION
Meloidae (bubble or oil bugs) have 177 species in Turkey and more than 3000 species in the world and cause great economic damage by feeding on the leaves and shoots of many fruits and vegetables [1-3].
The family Meloidae has been the subject of numerous taxonomic, biological, systematic, and faunistic studies and has received great attention worldwide [2]. LM and SEM were used in this study to examine the morphology of the male reproductive system of Mylabris cernyi (Meloidae), as there have been few investigations on the reproductive system of Meloidae [4].
A pair of testes, a pair of vasa deferentia, three pairs of accessory glands, an ejaculatory duct, and an aedeagus are the main organs of the male reproductive system in Meloidae [4]. The testis consists of many follicles, the number of which varies according to the species, where sperm development occurs. Accessory glands form the spermatophore, which allows the sperm to move and feed, and at the same time ensures the safe transport of the sperm to the female. The testis peritoneal sheath color, the testis shape, the testis follicle numbers, and the accessory gland shape and number vary interspecifically [4-9].
This study looked at the morphology of M. cernyi's male reproductive system to learn more about blister beetles in general.
MATERIALS AND METHODS
M. cernyi (n = 20) were collected in July 2019 from Keklikdere, Bingöl, Türkiye. The adults' organs were dissected under a stereo microscope (SM) after they had been sedated with ethyl acetate steam. After being moved to formalin, certain organs were embedded in paraffin. For light microscope (LM), slices (5–6 μm) were stained with hematoxylin and eosin (H and E). The other organs were transferred HMDS (Hexamethyldisilazane) and then dried with air. The organs were examined with a scanning electron microscope (JEOL JSM 6060 LV).
RESULTS AND DISCUSSION
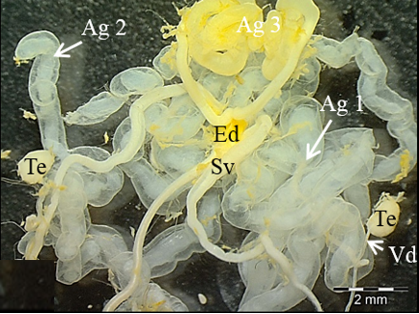
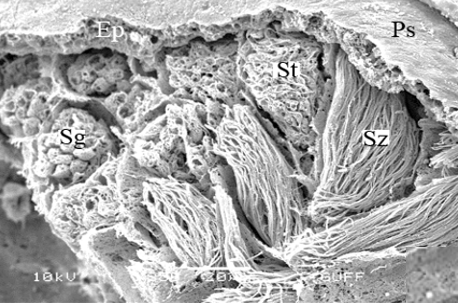
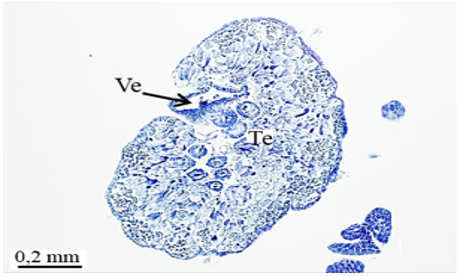
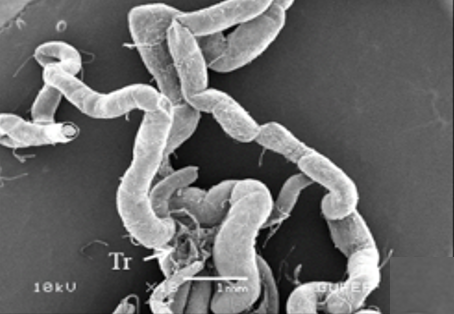
The male reproductive system of M. cernyi comprises a pair of testes (Te), a pair of vasa deferentia, a pair of seminal vesicles (Sv), three pairs of accessory glands (Ag 1- first pair of accessory glands, Ag 2- second pair of accessory gland, Ag 3- third pair of accessory gland), an ejaculatory duct (Ed) (Figure 1a), and an aedeagus (Ae) as in many other blister beetle species [4].
The testis
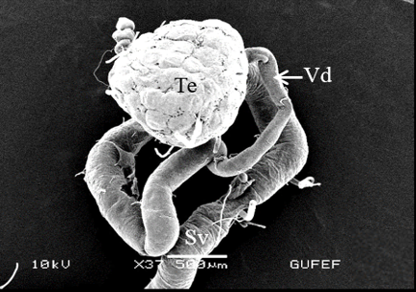
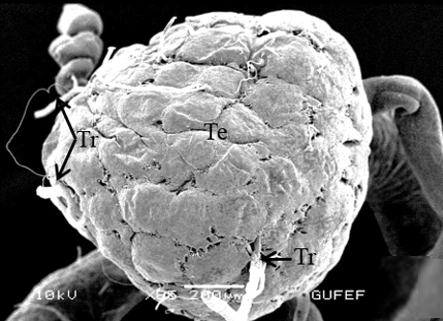
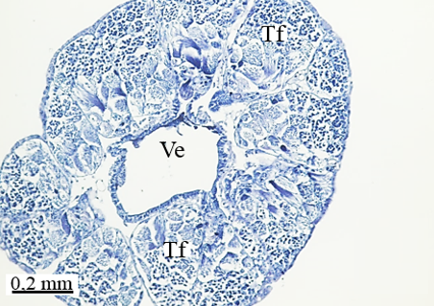
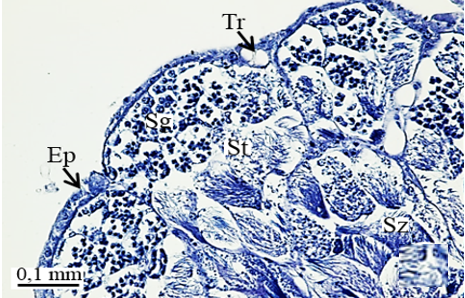
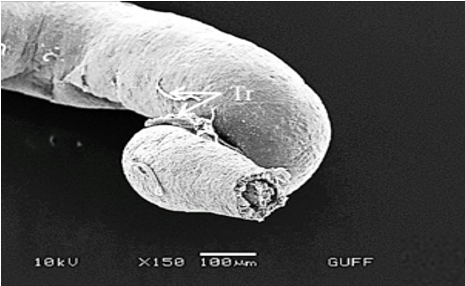
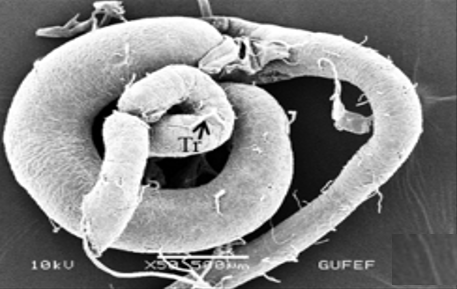
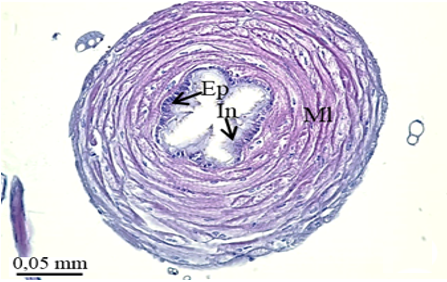
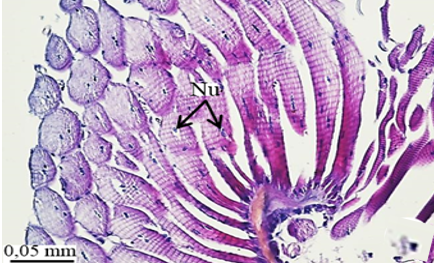
The testis of M. cernyi has a light yellow-colored (Figure 1a) globule-like structure (Figures 1b and 1c). There is a trachea (Tr) in some places on the testis surface (Figure 1c). The testis of M. cernyi is spherical (Figures 1a-1c) and the testis consists of testicular follicles (Tf), which have a radial arrangement. The size of the testis and the follicle number of tests may differ interspecifically. The testis of M. cernyi is ~12 mm (Figure 1c) and has 14–15 follicles (Figure 1d). Each testis of Meloe proscarabaeus Linnaeus, 1758 (Meloidae) contains 100–120 follicles [4].
|
|
|
a) |
|
|
|
b) |
|
|
|
c) |
|
|
|
d) |
|
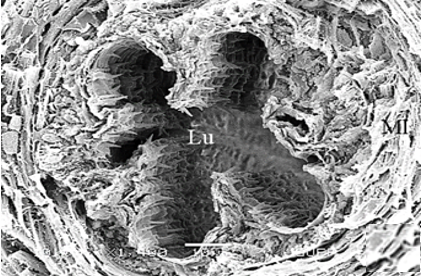
Figure 1. a) The male reproductive system in M. cernyi (SM). b) The testis, vas deferens, and seminal vesicle (SEM). c) SEM photograph of the testis which has a globule-like structure. d) The testis cross section (LM). |
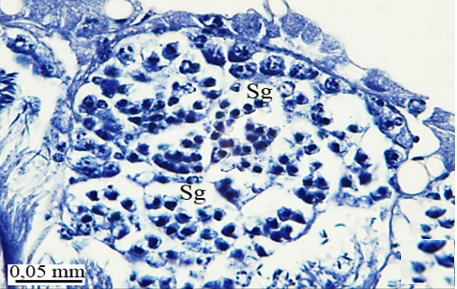
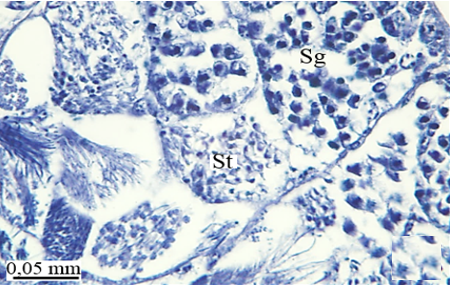
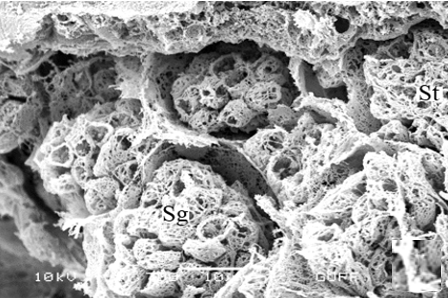
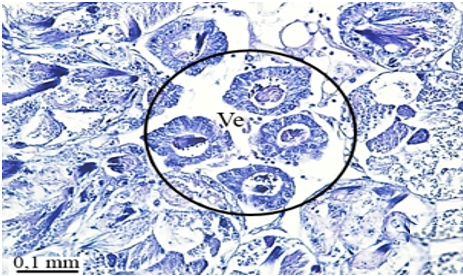
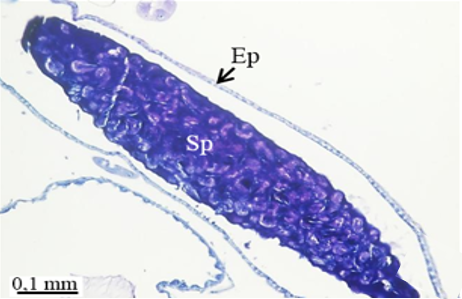
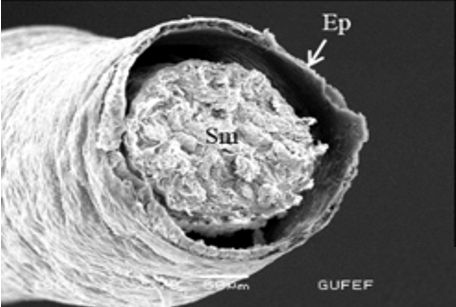
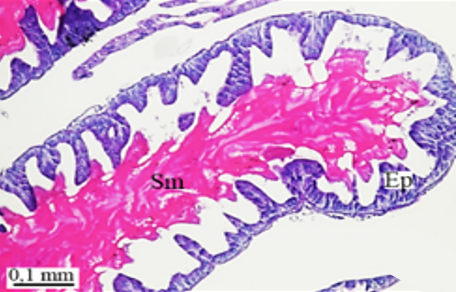
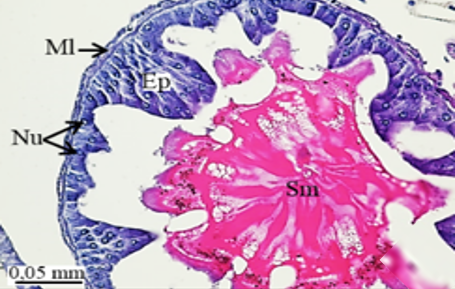
In testicular follicles of M. cernyi, 3 various zones (growth, maturation, and differentiation) are distinguished from the periphery of the testis to its center (Figures 1d; Figures 2a and 2b) as in Chrysolina herbacea (Duftschmid, 1825) (Chrysomelidae) and Chrysomela populi Linnaeus, 1758 (Chrysomelidae) [8, 9]. In the LM and SEM photographs of the testis section, it is seen that it is occupied by a monolayered epithelium (Ep) and peritoneal sheath (Ps) (Figures 2a and 2b).
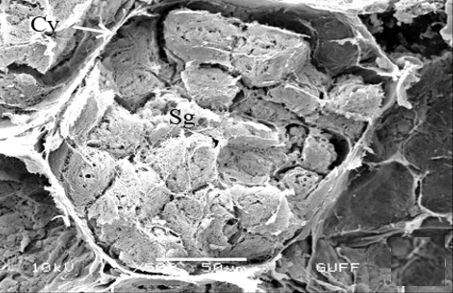
In the direction of the testis's edge, the growing zone is identifiable. Spermatocytes (Sg) within the cysts (Cy) may be identified in this instance (Figures 2c and 2d). The nuclei of spermatocytes are basophilic. Nearly the whole sperm head is covered with the nucleus (Figure 2c).
|
|
|
a) |
|
|
|
b) |
|
|
|
c) |
|
|
|
d) |
|
|
|
e) |
|
|
|
f) |
|
Figure 2. a, b) LM and SEM photographs of spermatocytes, spermatids, and spermatozoa in testicular follicles (LM, SEM), c, d) The spermatocytes in a cyst in the growth zone (LM, SEM), and e, f) The differentiation of spermatocytes into spermatids in the maturation zone (LM, SEM). |
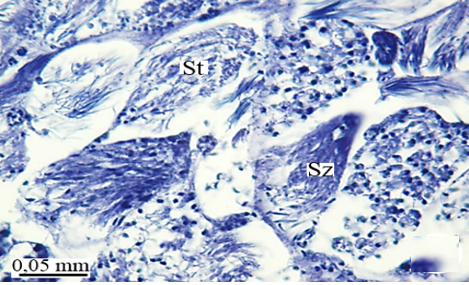
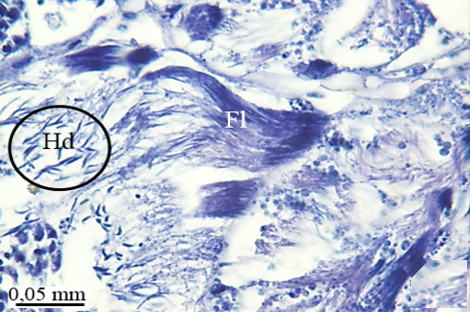
In the maturation zone, it is seen that the spermatocytes undergo meiosis, their structure completely changes, and they turn into spermatids (St) (Figures 2e and 2f). At this stage, the head and flagellum (Fl) of the spermatids are prominent. There is no regular sequence. The flagellum is made up of long, thin threads (Figures 3a and 3b).
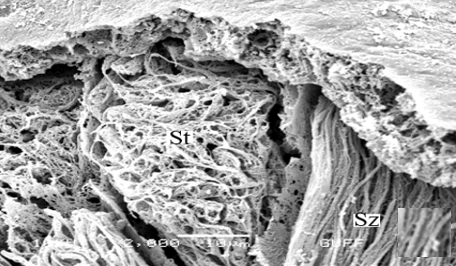
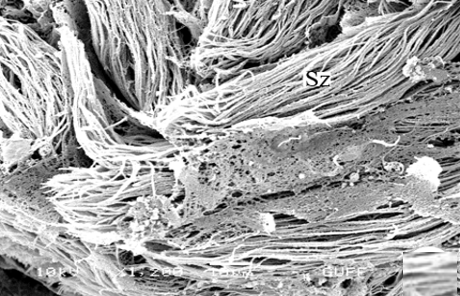
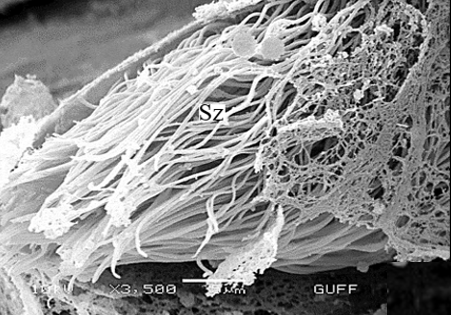
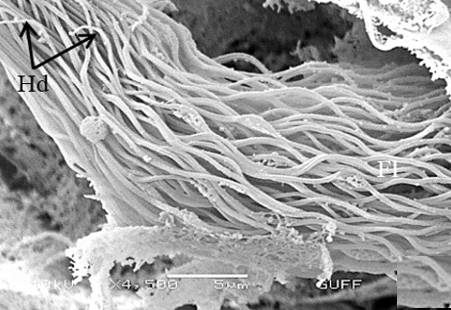
When approaching the vas deferens toward the testis center, it is seen that spermatids differentiate into spermatozoa (Sz) in the differentiation zone (Figures 3a and 3b). It can be distinguished that spermatozoa are in regular bundles (Figures 3a-3f). In the LM and SEM photos, it is seen that the sperm heads (Hd) are quite thin and show the appearance of a hook (Figures 3e and 3f).
|
|
|
a) |
|
|
|
b) |
|
|
|
c) |
|
|
|
d) |
|
|
|
e) |
|
|
|
f) |
|
Figure 3. a, b) LM and SEM photographs of the transformation of spermatids into spermatozoa in the differentiation zone (LM, SEM), c-f) The regular spermatozoa bundles (LM, SEM). |
The vas efferens, vas deferens, and seminal vesicle
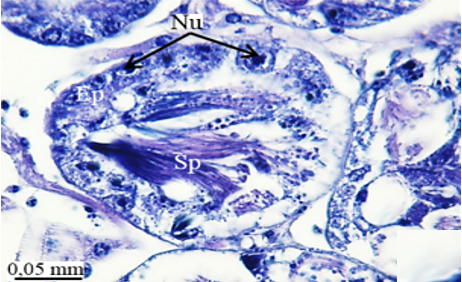
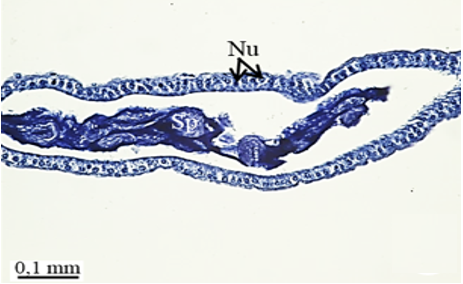
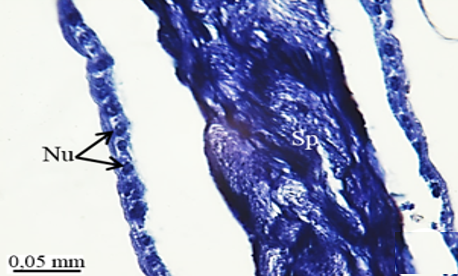
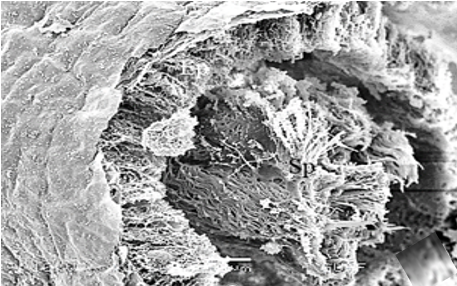
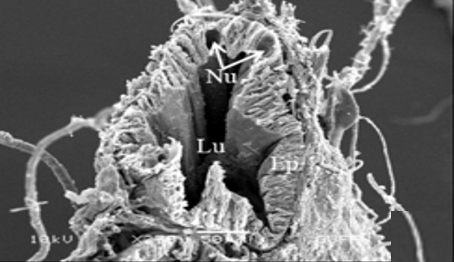
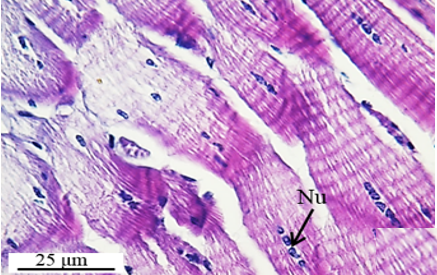
The testis emerges as a vas efferens (Ve) (Figures 4a and 4b) that later joins forming a vasa deferentia. In the LM and SEM photographs, mature sperm (Sp) masses are distinguished in the lumen (Lu) of the vas efferens (Figures 4b-4d). The vas efferens wall is surrounded by monolayer epithelium. Epithelial cells have round and basophilic nuclei (Nu) (Figure 4c).
|
|
|
a) |
|
|
|
b) |
|
|
|
c) |
|
|
|
d) |
|
|
|
e) |
|
|
|
f) |
|
|
|
g) |
|
|
|
h) |
|
|
|
i) |
|
|
|
j) |
|
Figure 4. a, b) The histological section of testis and vas efferens, c, d) LM and SEM photographs of vas efferens, e-h) LM and SEM photographs of vas deferens, i, j) LM and SEM photographs of seminal vesicle. |
Like in C. herbacea and C. populi (Chrysomelidae), the vas efferens carry the adult sperm masses from testis follicles to the vas deferens [8, 9].
Vas deferens is rather narrow and near the testis. The seminal vesicle is narrow (Figure 1b). However, the histological structure of the vas deferens and seminal vesicle is the same (Figures 4e, 4g, and 4i). Both are surrounded by a simple cuboidal epithelium (Figures 4e-4j) as in Pimelia subglobosa (Pallas, 1781) (Tenebrionidae) [10]. The vas deferens wall of Capnodis tenebrionis (Linnaeus, 1761) (Buprestidae) has a simple cylindrical epithelium [5].
Vas deferens and seminal vesicle epithelium in M. cernyi have round nuclei in the middle of the epithelial cell (Figures 4g and 4i). It is distinguished in their lumen that mature sperm bundles are stored (Figures 4e-4j) as in C. tenebrionis (Buprestidae), C. herbacea (Chrysomelidae), and C. populi (Chrysomelidae) [5, 8, 9]. The vas deferens are connected to the ejaculatory duct via a seminal vesicle.
The accessory glands
In M. cernyi, there are three pairs of accessory reproductive glands (Figure 1a) as in M. proscarabaeus (Meloidae) [4].
The number and shape of accessory glands vary interspecifically. In C. herbacea (Chrysomelidae), there are two tubular accessory glands [8]. Tanymecus dilaticollis Gyllenhal, 1834 (Curculionidae) has two accessory glands (blind-ended tubules) and two prostate glands (multilobed) [7].
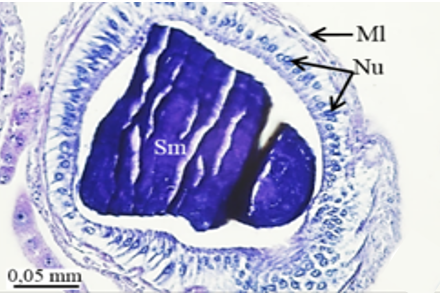
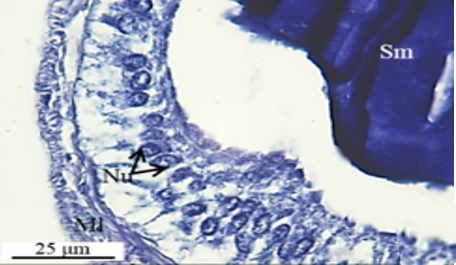
The accessory glands open into an ejaculatory duct (Figure 1a). The first pair of accessory glands of M. cernyi is a pair of short, tubular, and light yellow (Figure 1a). It has long columnar epithelial cells and a muscle layer (Ml) (Figures 5a-5d). Its lumen contains basophilic secretory materials (Figures 5a and 5c).
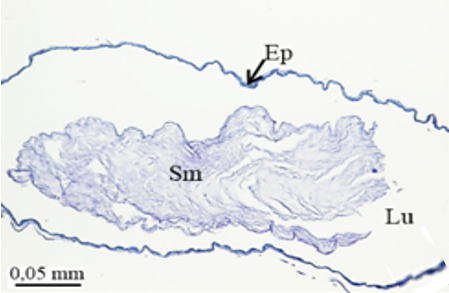
The second pair of accessory glands of M. cernyi is long, coiled, tubular, and whitish (Figure 1a). They have basophilic and fibrous secretory materials (Sm) in their lumen (Figures 5e and 5f). The wall of this gland is surrounded by monolayered squamous epithelium (Figures 5e, 5f, and 5h). The trachea is distinguished in some places on the surface of the second pair of accessory glands in the SEM photograph (Figure 5g).
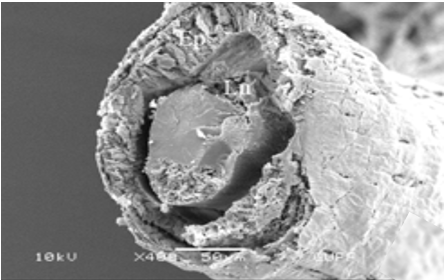
The third pair of the accessory glands of M. cernyi is long, coiled, and tubular (Figures 1a and 5l). Its lumen contains eosinophilic secretory materials (Figures 5i and 5j). The trachea is distinguished in some places on the surface of the third pair of accessory glands in SEM photographs (Figure 5k). The wall of this gland has a pseudostratified epithelium with oval nuclei and muscle. The epithelium is recessed into the lumen (Figures 5i, 5j, and 5l).
|
|
|
a) |
|
|
|
b) |
|
|
|
c) |
|
|
|
d) |
|
|
|
e) |
|
|
|
f) |
|
|
|
g) |
|
|
|
h) |
|
|
|
i) |
|
|
|
j) |
|
|
|
k) |
|
|
|
l) |
|
Figure 5. a-d) LM and SEM photographs of a cross-section of the first pair of accessory glands at different magnifications, e, f) The longitudinal sections of the second pair of accessory glands, g) SEM photographs of the second pair of accessory glands, h) SEM photographs of the cross-section of the second pair of accessory glands, i, j) The longitudinal and cross sections of the third pair of accessory glands (LM), k) SEM photograph of the third pair of the accessory gland, l) SEM photograph of cross section of the third pair of accessory gland. |
The ejaculatory duct
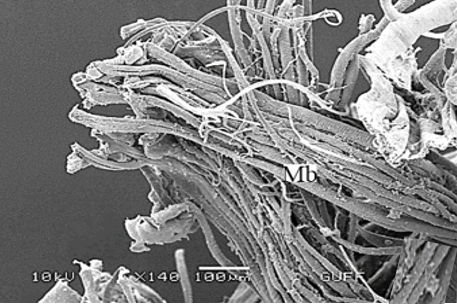
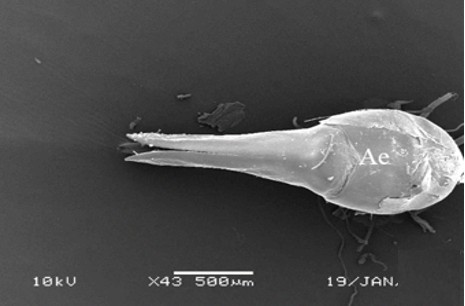
The wall of the ejaculatory duct is surrounded by the intima (In), epithelium, and a thick muscle layer, as in other species [5-7, 10]. The epithelium has deep recesses toward the lumen (Figures 6a and 6b). There are spines (Spi) on the lumen facing the intima layer surface (Figures 6c and 6d). Histological sections show multinucleated striated muscle bundles (Mb) between the ejaculatory duct and the aedeagus (Figures 7a-7c). The ejaculatory duct opens out with the aedeagus structure. In SEM photographs, it is observed that the aedeagus has a flat surface (Figure 7d).
|
|
|
a) |
|
|
|
b) |
|
|
|
c) |
|
|
|
d) |
|
Figure 6. a, b) LM and SEM photographs of a cross-section of the ejaculatory duct, c, d) SEM photographs of spines extending from the intima of the ejaculatory duct. |
|
|
|
a) |
|
|
|
b) |
|
|
|
c) |
|
|
|
d) |
|
Figure 7. a, b) The histological section of striated muscles between the ejaculatory duct and aedeagus (LM), c) SEM photograph of muscle bundles between the ejaculatory duct and aedeagus, d) SEM photograph of aedeagus. |
CONCLUSION
The morphology of M. cernyi species' male reproductive system is described in this paper. This work advances our understanding of the reproductive organs of the Coleoptera order, which includes the Meloidae. Additionally, the testis color, shape, follicle number, and accessory gland shape and number may differ interspecifically. Therefore, this study also contributes to systematic studies.
ACKNOWLEDGMENTS: The authors would like to thank Gazi University Academic Writing Application and Research Center for proofreading the article.
CONFLICT OF INTEREST: None
FINANCIAL SUPPORT: None
ETHICS STATEMENT: None