Entomology and
Applied Science Letters
Applied Science Letters
 2023
Volume 10
Issue 1
2023
Volume 10
Issue 1

The present studies on the alimentary canal of the last instar larva of Deudorix isocrates (Fab.) describe the anatomical and histological details which have not been reported earlier. The straight alimentary canal is divisible into foregut: buccal cavity, pharynx, esophagus, and crop; midgut and hindgut: pylorus, ileum, colon, and rectum. The wall of the buccal cavity, pharynx, and esophagus is composed of cuboidal epithelium which is produced into six longitudinal folds. The intima of the posterior pharynx is characteristic because it shows longitudinal overlapping ridges and backwardly directed bristles. The product has thin intima and numerously folded squamous epithelium. Musculature is very thin in this region. The stomodaeal valve is derived from the foregut with characteristic histological features. The anterior and posterior interstitial rings are composed of small columnar epithelium. The midgut wall is composed of simple columnar epithelium with scattered goblet cells and individually distributed regenerative cells, the luminal side has the peritrophic membrane, and the Midgut musculature is thin and consists of an inner thin circular muscle layer and outer longitudinal muscle bundles. The epithelium of the hindgut in the pylorus colon and rectum is cuboidal, while in the ileum it is squamous type. The intima of the hindgut is relatively thin compared to the foregut but it produces patches of spines at the posterior pylorus. The rectum forms the cryptonephridial system.
INTRODUCTION
Deudorix isocrates (Fab.) is a serious economic pest on pomegranate and guava [1, 2], commonly called pomegranate fruit borer. It is the main constraint in producing adequate quantities and quality pomegranate fruits. The pomegranate fruit borer is the most common, polyphagous, and damaging pest, with a wide range of host plants including pomegranate, citrus, guava, litchi, apple, peach, pear, plum, and sapota [3]. It has received much attention regarding its bio-ecological studies and management strategies to minimize its attack on pomegranate crops. No anatomical or histological work has been done, however, these studies have great importance in the field of pest control. The anatomy, histology, and physiology of the alimentary canal of arthropods are related to the digestion and absorption of their specific diet [4]. The midgut, which occupies the largest part of the alimentary canal, plays a major role in the digestion of food and absorption of nutrients, but also chemical and biological insecticides [5-9]. Due to this, the anatomical and histological studies have greater significance in assessing the efficacy of plant extract with insecticidal properties [10-13], evaluation of the pathogenicity of biological control agents [14], the toxicity of synthetic insecticides [15], the effect of drugs [16] on lepidopteran pests, leading to an increase of interest for knowledge about anatomy and histology of the alimentary canal of the insect pest species. Our study is the first detailed anatomical and histological description of the alimentary canal of a D. isocrates larva. The present study provides the fundamental anatomical and histological data of the alimentary canal of D. isocrates which is useful in toxicological and pathological research on this pest.
MATERIALS AND METHODS
The specimens were obtained by laboratory rearing as well as from the pomegranate filed by collecting infested fruits due to D. isocrates at larval stages. For the anatomical study, the fresh larvae were sacrificed by putting them in hot water. The sacrificed specimens were opened along with the mid-dorsal line in insect saline under a stereoscopic dissecting binocular microscope. The morphometric observations were made using ocular and stage micromere. These observations were presented as average with SE (Standard Error) (Table 1).
Table 1. Morphometric observations on the alimentary canal of D. isocrates larva.
|
Organs of the alimentary canal |
Average Length (± SE) |
Average Width (± SE) |
|
Pharynx |
0.742 (± 0.041) |
0.631 (± 0.031) |
|
Oesophagus |
0.902 (± 0.056) |
0.578 (± 0.014) |
|
Crop |
0.578 (± 0.022) |
1.173 (± 0.065) |
|
Midgut |
16.780 (± 0.887) |
3.976 (± 0.262) |
|
pylorus |
2.020 (± 0.092) |
1.810 (± 0.043) |
|
Ileum |
3.470 (± 0.247) |
2.280 (± 0.128) |
|
Rectum |
2.800 (± 0.213) |
2.280 (± 0.090) |
|
Total |
27.294 (± 1.105) |
|
Note SE: Standard Error
The live specimens from laboratory-reared stock were also used to make histological preparations. The specimens were dissected in 0.7% saline [17]. After dissecting the complete alimentary canal, it was cut into six pieces: one with foregut and a small portion of the midgut, four pieces of the midgut, and one with hindgut and a small portion of the midgut. The midgut peritrophic membrane with luminal content was removed before fixing the tissue. The pieces of the alimentary canal were immediately transferred to different vials containing fixatives like aqueous Bouin’s, Carnoy’s, formal saline, and chilled Acetone. Out of these fixatives, Bouin’s was the best and hence it was used for further histological studies. The organs were kept in Bouin’s for 12 hrs., then washed under running tap water for 5 to 6 hrs., dehydrated in ascending grades of alcohol, cleared in Benzene, and infiltrated with paraffin wax (M. P. 52–54 oC) for two and half hours. The longitudinal serial sections and cross serial sections were cut at 7 µ with a rotary microtome. Sections were spread on pre-egg albumin-smeared slides [18].
The staining of tissue sections was done with Delafield’s hematoxylin and 1% alcoholic eosin for the histological studies. The stained sections were mounted in DPX after clearing in xylene. The histological observations were made using a light microscope under 10X and 45X magnifications.
RESULTS AND DISCUSSION
The alimentary canal of the D. isocrates larva is almost straight and more or less equal to the body length. Anatomically it is well distinguished into foregut; stomodaeum, midgut; ventriculus, and hindgut; Proctodaeum. These demarcations can be determined by the dilations and constrictions at appropriated points along the length of the alimentary canal. Histologically the wall of the alimentary canal is typically composed of a simple epithelium surrounded by bundles of longitudinal muscles and a layer of circular muscles. The cells of the foregut and hindgut, both of ectodermal origin; secrete cuticular intimal lining the lumen of the alimentary canal [19].
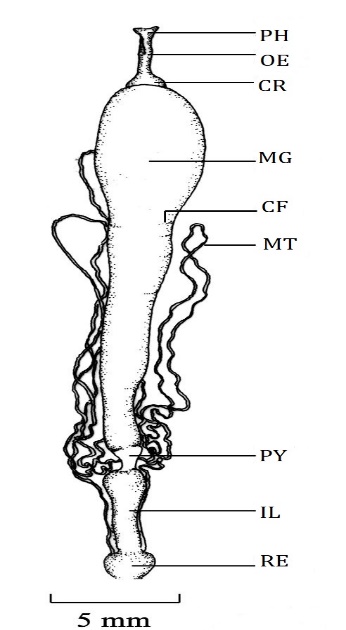
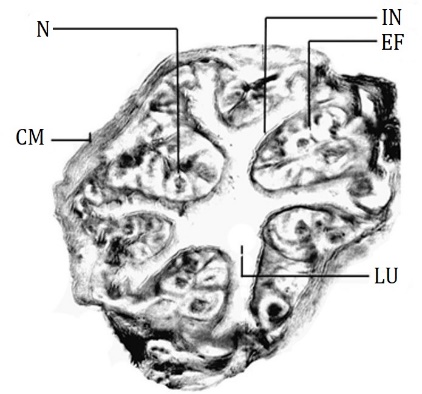
The foregut is divisible into four regions buccal cavity, Pharynx, esophagus, and crop [20] (Figure 1) as the general anatomical feature of the lepidopteran larva. The buccal cavity is the anteriormost part of the foregut [8]. Histologically the wall of the buccal cavity is composed of thin intima, very thin squamous epithelium, a thin layer of circular muscles, and bundles of longitudinal muscles which are situated inside folds. The pharynx lies anterior to the supra oesophageal ganglia and joins with the esophagus which lies posterior to the supra oesophageal ganglia. The pharynx is divided into two regions; the anterior pharynx and the posterior pharynx. The Intima of the anterior pharynx is thicker than that of the buccal cavity. Epithelial lining is composed of cuboidal cells having centrally placed round nuclei. The lining is thrown into six longitudinal folds and it is surrounded by a thick layer of circular muscles (Figure 2). Wherever folds are broad the bundles of longitudinal muscles are seen below the epithelial lining. The posterior pharynx shows thick intimal lining which is produced into overlapping ridges with backwardly directed bristles while cuboidal epithelium and circular muscle layer are similar to that of the anterior pharynx.
|
|
|
Figure 1. Alimentary canal of D. isocrates larva CR: Crop, IL: Ileum, MG: Midgut, MT: Malpighian tubule, OE: Oesophagus, PH: Pharynx, PY: Pylorus, RE: Rectum. |
|
|
|
Figure 2. T.S. of the anterior pharynx, D. isocrates larva x113 CM: Circular muscle, EF: Epithelial fold, IN: Intima, LU: Lumen, N: Nucleus. |
The pharynx opens into to narrow passage like the esophagus. The intimal lining of the esophagus is thinner than that of the pharynx and it does not bear ridges or bristles. The epithelium is composed of a bit large cuboidal cells with compactly arranged oval nuclei. The cuboidal epithelium of the esophagus is present in other lepidopteran species like Heliothis virescense [21]. The layer of circular muscles is comparatively thin. The longitudinal muscles run through epithelial folds and the circular muscle layer encircles the epithelial lining.
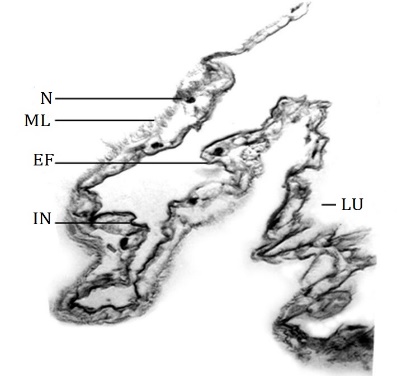
The esophagus extends up to the mid-region of the prothoracic segment where it widens into a saclike structure, the crop [22, 23]. It was considered a simple dilatation of the posterior region of the esophagus [24]. The crop is thin-walled and translucent. The functions of the crop are the storage and flow of ingested food [24, 25]. The wall of the crop is composed of thin intimal lining and deeply folded thin squamous epithelium (Figure 3) which permits a large increase in volume to store food [4]. The muscle layer is difficult to locate from the basement membrane of the epithelium at certain places. This observation is similar to that in the larva of Lymantria dispar [4].
|
|
|
Figure 3. T.S. of the crop – in part, D. isocrates larva x125 EF: Epithelial fold, IN: Intima, LU: Lumen, ML: Muscle layer, N: Nucleus of squamous epithelial cell. |
The crop leads into the midgut region. At the site of the junction between the foregut and hindgut, the ventral wall of the midgut extends much more anteriorwards than the dorsal side. Thus it forms an oblique connection. The crop appears to be pushed posteriorly towards the mid-gut. Internally, the end of the crop is projected in the lumen of the midgut. This internal extension of the foregut forms the stomodaeal valve [22]; also called the cardiac valve [4] which is homologous to the proventriculus [26].
After a short distance from the base, the stomodaeal valve does not make a complete cylinder. About half of the part of the cylinder runs much posteriorly than the remaining portion. The lateral edges of the much-extended region are curved inwardly and try to form a cylindrical passage at the posterior-most region of the stomodaeal valve. Histologically it bears two layers, adaxial and abaxial similar to that reported in H. armigera [23]. The adaxial layer is the inner one, which runs in the midgut for a short distance and then turns anteriorwards as the abaxial layer. Hence, the epithelial lining of the two layers lies opposite but close to each other. The intima of the stomodaeal valve is comparatively thicker than that of the crop. The abaxial layer ultimately joins with the wall of the midgut where they meet each other via the anterior interstitial ring. At the junction between the stomodaeal valve and interstitial ring, the intima is remarkably thick and forms a collar-like structure (Figure 4) while the interstitial cell lining is lined internally by thin intimal lining. The epithelium is composed of simple columnar cells with oval nuclei. The foregut musculature comprises outer circular and inner longitudinal muscles [27]. This arrangement is prevalent in lepidopterist larvae of different species.
|
|
|
Figure 4. Magnified L.S. passing through the anterior interstitial ring, D. isocrates larva x443. AIR: Anterior interstitial ring, INC: Collar formed by thick intima, IN: Intima, LUM: Lumen of the midgut, N: Nucleus of columnar epithelial cell. |
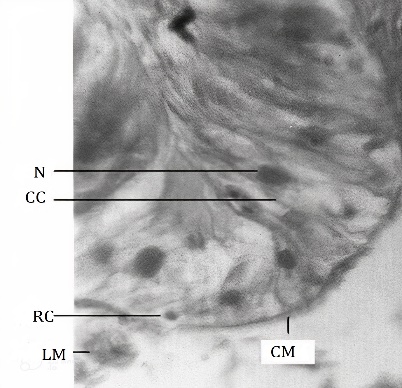
The midgut is longer than other parts of the alimentary canal (Figure 1). It extends from the mesothoracic segment up to the 4th segment of the abdomen. It is held in position by tracheal attachments. Its anterior half is relatively distended than that of the posterior half. The wall of the midgut is thicker than that of the foregut and the hindgut. The wall bears crosswise folds when it is devoid of luminal contents. These foldings reported in the gut wall of lepidopteran species give elasticity to the gut wall which allows a large amount of food to pass through it [21]. The wall bears several small buds like gastric caeca throughout its length. The gastric caeca are not visible externally due to the crosswise folds of the midgut wall. The wall of the midgut is composed of the peritrophic membrane, epithelium, and muscle layers. The peritrophic membrane acts as a barrier to prevent food particles from coming into contact with the microvilli of the midgut cells, thereby avoiding damage to the cells [28]. In this study, the peritrophic membrane has been taken off along with the gut contents to achieve good fixation. Hence, the peritrophic membrane is not seen in the sections studied. The midgut epithelium (Figure 5) consists of simple columnar epithelial cells, regenerative and goblet cells as reported in the larva of most of the lepidopteran species [29-31]. The simple columnar cells are tall with compactly arranged ovoid to round nuclei. The nuclei are situated misally or apically. The regenerative cells are distributed individually at the base of epithelium along the basement membrane unlike that present in Lymantria dispar larva [4] but similar to that present in H. cecropia [19]. This type of distribution of regenerative cells is the expected feature of a continuous feeder. The regenerative cells in the insects help in epithelium renewal and facilitate gut growth during the ecdysis [32-34]. The mesenteric caeca are distributed throughout the length of the midgut. The epithelium at the mesenteric caeca is similar to that of the midgut epithelium. The circular muscle layer is thin. Bundles of longitudinal muscles are scattered around the circular muscle layer.
|
|
|
Figure 5. Magnified part, T.S. of the midgut, D. isocrates larva x375 CC: Columnar epithelial cell, CM: Circular muscle, LM: Longitudinal muscle, N: Nucleus, RC: Regenerative cell. |
The hindgut of the D. isocrates larva is distinguished into the pylorus, ileum, colon, and rectum (Figure 1) like most of the lepidopteran species [7, 19, 21, 35]. Histologically, the hindgut begins with the posterior interstitial ring. The intimal lining of this region is thick. At the posterior end of the midgut, the midgut epithelium is continued with the epithelium of the posterior interstitial ring. The epithelium is composed of simple columnar cells having oval nuclei similar to that of Hieliothis virescense [21] However, in some species like Anticarsia gemmatalis and Heliothis zea the epithelium of the posterior interstitial ring is composed of cuboidal cells [7, 36]. Like the anterior interstitial ring, the cells have little affinity with the cytoplasmic and nuclear stains, i.e. eosin and hematoxylin.
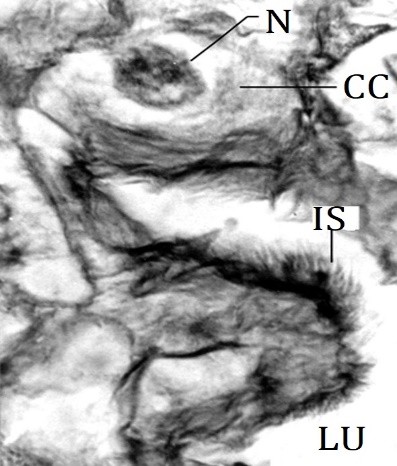
The pylorus occupies the 5th abdominal segment. The pyloric region appears to be pushed anteriorly in the posterior end of the midgut. As compared to the midgut, the wall of the pylorus is thin and without crosswise folds. The anterior portion of the pylorus is wider. It gradually narrows posteriorly to give rise tubular part. Thus, in general, the pylorus is a funnel-shaped region. A pair of ducts arise from the lateral sides of the posterior part of the pylorus and divides each into three branches. The branching pattern is peculiar. From the common duct, two branches arise and then out of two branches, one branch bifurcates while the other branch remains unbranched. Thus, all 3 branches are held together on either side of the pylorus. These branches are the Malpighian tubules. The distal part of Malpighian tubules runs anteriorwards along the wall of the midgut up to the 2nd abdominal segment where they turn posteriorwards up to the rectum. They enter the wall of the rectum and thus represent cryptonephridial condition. Similar observations have been made in budworm, Heliothis virescense [21]. The pylorus is histologically distinguished into a pyloric valve, anterior pylorus, and posterior pylorus. The posterior interstitial ring and the anteriormost region of the pylorus collectively form the inwardly projecting fold-like pyloric valve. The intima of the pyloric valve is thick. The cuboidal epithelium of the pyloric valve is composed of large cuboidal cells and a few scattered small cuboidal cells. The intima of the anterior pylorus is thin as compared to the intima of the pyloric valve. The epithelium is composed of large cuboidal cells with a large round misally placed nucleus. The epithelial folds of the anterior pylorus are small and irregular. The intima of the posterior pylorus is remarkably different as compared to that of the anterior pylorus. It is produced into patches of sclerotized spines (Figure 6) in some regions. The spines on the hindgut intima are to assist in breaking off the peritrophic membrane after it exits from the midgut [37] and these also might assist in propelling the feces [4]. The epithelium is composed of large cuboidal cells with misally placed round nuclei. The epithelium of the posterior pylorus is thrown into very conspicuous irregular folds. The conspicuous folds give rise to a thick wall at the posterior pylorus and these folds remarkably occlude the lumen at the opening into the ileum. At the junction of the posterior pylorus and the ileum, the epithelium develops into longitudinal folds. These folds run anteroposteriorly and continue with the epithelial folds of the ileum. The circular muscle layer surrounds the epithelium. The longitudinal muscle bundles lie inward and very close to the base of epithelial folds.
|
|
|
Figure 6. Magnified part of epithelial fold from posterior pylorus D. isocrates larva x540 CC: Cuboidal epithelial cell, LU: Lumen, N: Nucleus, IS: Patch of intimal spines. |
The pylorus opens into the ileum. The ileum is the longest part of the hindgut. It lies in the 6th and 7th abdominal segments. Anteriorly the ileum is wider and posterior wards it narrows gradually. The intima of the ileum is wavy. The epithelium is composed of squamous epithelial cells similar to that of Heliothis virescense [21]. The nucleus of squamous cells is spherical. The epithelium is thrown into numerous longitudinal folds. The muscle layer comprises the inner thick circular muscle layer and outer bundles of longitudinal muscles.
The ileum opens into the rectum through an anatomically indistinct colon. The colon is indistinct externally in other species of Lepidoptera like Heliothis virescense [21]. The Intima of the colon is also wavy like the intima of the ileum. The epithelium is composed of cuboidal cells having a spherical nucleus. The nucleus is placed misally or basally. The epithelium is peculiarly thrown into complex folds, which occlude the lumen. The muscle layer is relatively thick. It consists of an inner thick circular muscle layer and outer bundles of longitudinal muscles. The rectum is a roughly round sac with a thin and translucent wall. Rectum opens outside by an anal opening, which is on the ventral side of the 10th segment of the abdomen. The wall of the rectum is traversed by Malpighian tubules and reveals cryptonephridial condition as it is typically found in Lepidoptera larvae [38]. The Intima of the rectum is thin. The epithelium is composed of large cuboidal cells with round nuclei. The epithelium is thrown into folds having irregular shapes and sizes. The muscle layer consists of inner thick circular muscles and outer longitudinal muscle bundles.
Results should be clear and concise. This should explore the significance of the results of the work, not repeat them. A combined Results and Discussion section is often appropriate. Avoid extensive citations and discussion of published literature.
CONCLUSION
In D. isocrates larva the straight alimentary canal with anatomically well-distinguished foregut, midgut and hindgut is more or less equal to the body length. Histologically the wall of the alimentary canal is typically composed of a simple epithelium surrounded by bundles of longitudinal muscles and a layer of circular muscles. The epithelium of the foregut and hindgut is lined with cuticular intima while the peritrophic membrane is present in the midgut; relatively the intima of the foregut is thicker than that of the hindgut. The foregut is divisible into four regions buccal cavity, Pharynx, esophagus, and crop. Internally the crop develops the stomodaeal valve which extends into the midgut lumen. The intima of the posterior pharynx is characteristically produced into overlapping ridges and backwardly directed bristles while it is smooth and thin in the buccal cavity, esophagus, and crop. The numerously folded thin squamous epithelium is present in the crop while in other regions of the foregut, it is cuboidal and produced into six longitudinal folds. The circular muscles encircle the epithelium and longitudinal muscle bundles are visible through the bases of epithelial folds up to the oesophageal region. In the crop, the musculature is very thin and covers the epithelium externally. The midgut is the longest region of the alimentary canal. It is composed of an inner peritrophic membrane, simple columnar epithelium with goblet cells, and individually distributed regenerative cells towards the basement membrane. Gastric caeca has similar histological features to that of the midgut wall. Histologically, the midgut is delimited by anterior and posterior interstitial rings which are composed of small columnar epithelium. The hindgut constitutes the pylorus, ileum, colon, and rectum with a cryptonephridial system of excretion. The intima of the hindgut is relatively thinner than that of the foregut. It is produced into patches of sclerotized spines at the posterior pylorus. The cuboidal epithelium of pylorus produces longitudinal conspicuous folds which occlude the lumen. The squamous epithelium of the ileum is thrown into numerous longitudinal folds lined with wavy intima. The muscle layer comprises the inner thick circular muscle layer and outer bundles of longitudinal muscles. In the colon, the epithelium is cuboidal and thrown into numerous complex folds. The cuboidal epithelium of the rectal wall also produced folds with irregular shapes and sizes. The thick musculature constitutes inner circular and outer longitudinal muscle bundles. The present work on the alimentary canal provides a detailed description of anatomy and histology which is important in further research on the physiology, assessment of the toxicity, and pathogenicity of various herbal and synthetic insecticides against this pest and useful to design pest management strategies against this pest.
ACKNOWLEDGMENTS: The author is grateful to Dr. P.V. Joshi (Professor, Dept. of Zoology Savitribai Phule Pune University) for his sincere help and guidance.
CONFLICT OF INTEREST: None
FINANCIAL SUPPORT: None
ETHICS STATEMENT: The sampling and research work was performed according to the guidelines of the academic institute.