Entomology and
Applied Science Letters
Applied Science Letters
 2024
Volume 11
Issue 3
2024
Volume 11
Issue 3

In this study, the effect of the intracellular symbiotic bacterium Wolbachia on the search power, access time, and exit percentage of female (A), natural hermaphrodite (B), and antibiotic-treated hermaphrodite (CB) Trichogramma brassicae wasps was investigated in the laboratory and cold storage. The functional response test of these three populations to different densities of flour moth eggs was obtained from type II and no significant difference was observed between their search power (a) and access time (Th). Based on the results of short-term cold storage, the average total exit percentage of natural hermaphrodite, bacterially infected female, and antibiotic-treated hermaphrodite bees reared at 20 °C was significantly higher than that of 25 °C. However, during similar storage periods, all three populations were placed in common groups in terms of exit percentage without significant differences. According to the results of this study and previous findings, by providing optimal conditions for reproduction and maintenance of Wolbachia-infected lines, the use of unisexual bees can have a relative advantage compared to natural bisexual bees, which is discussed in this article.
INTRODUCTION
What is being discussed in agricultural management today is reducing the use of chemical pesticides, preserving the environment, supporting natural enemies, and ultimately producing healthy food. Therefore, biological control specialists are always looking for solutions to improve and increase the efficiency of natural enemies in different regions and ecosystems. Among the factors that ensure the success of a natural enemy before release is the recognition of measurable characteristics in a suitable population or ecotype [1-3]. Among these criteria, the sex ratio created by the female wasp is an important qualitative criterion, and since female parasitoid wasps are the main factor in killing the pest, hypotheses have been made regarding the advantage of using species or populations of single-sex parasitoid wasps in the release against a pest [4, 5].
Trichogramma wasp, with more than 180 species, is the largest and best-known genus of the Trichogrammatidae family. Due to its ease of mass reproduction and the possibility of using its various species as parasitoids of lepidopteran eggs that damage crops, it has the highest level of release in the world among natural enemies of pests in terms of application [6, 7]. After the introduction of female-producing species of Trichogramma wasp in the 1990s, many studies were conducted on the factors that cause and the advantages of using unisexual Trichogramma wasps [5]. The factor of unisexual reproduction of bees (thelytokous parthenogenesis) can have a genetic or microbial origin that causes the development of females from unfertilized eggs. In the Trichogramma cacociae (Marcal), the main cause of genetic female reproduction has been identified [8]. Wolbachia bacteria are known to induce parthenogenesis in more than 18 species of Trichogramma bees and there is also the possibility of horizontal transmission within and between species [9].
The results of numerous studies conducted to determine the different effects of the symbiotic bacteria Wolbachia on host fitness indicate very different effects of this bacterium on biological, olfactory, and morphological indices of Trichogramma bees, including fertility, lifespan, parasitism rate, exit percentage, dispersal power, host recognition, leg length, etc. Some of the results related to the topic will be discussed [10-16].
In this study, while adding a bisexual population resulting from the removal of bacteria from the body of infected bees using antibiotics and determining the functional response of three unisexual and bisexual populations of Trichogramma brassicae, the result of storing parasitized eggs of the mealybug was examined and compared by the aforementioned populations.
MATERIALS AND METHODS
Collection and identification of ecotypes of Trichogramma bee reproductive material
To obtain the reproductive population of Trichogramma bees, parasitic eggs were collected from weeds, including Xanthium stramarium L. (Asterales: Asteraceae), and corn and rice fields. After transferring the parasitic eggs to a constant temperature room and leaving the bees, each of the strains separately from the collection location and host type at a temperature of 60 ± 10 °C, a relative humidity of 60 ± 10%, and a period of 16 hours of light and 8 hours of darkness on the eggs. Sterilized Ephestia kuehniella (Lep.: Pyralidae) were reared, and each of them was placed individually in test tubes (12 × 75 mm) before the bee hatched from the emergence of female bees. Immediately after the emergence of female bees, paper strips containing fresh Ephestia eggs were provided to each of them, and the type of virginity in virgin bees was determined by the isofemale method.
By observing female reproductive individuals in each of the populations, a pure line was created and its breeding continued in appropriate and controlled conditions. To prove the possibility of female bees being produced by Wolbachia bacteria, first, female bees were fed with a solution of water, honey, and antibiotics [12]. And again, their reproduction method was checked after 3-4 generations. In case of returning to the bisexual state, there is a high possibility of infection with bacteria-inducing virginity, and for the final confirmation, the molecular method and the specific primer of the wsp gene were used [17]. To identify bees, in addition to using the key related to morphological traits, the molecular identification method based on the ITS2 region was also used [18].
Creation of bisexual and reproductive populations infected with Wolbachia bacteria
In this research, to evaluate the effect of Wolbachia bacteria and compare some characteristics of bisexual and female populations, three purified Trichogramma brassicae lines with the abbreviations A (Asexual), B (Bisexual), and CB (Cured Bisexual) were created by single material method. Line A was infected with PI-Wolbachia. In population B, there was bisexual reproduction and the bisexual line CB was created using antibiotic treatment for four generations of female bees infected with the bacteria (A) with water and honey impregnated with the antibiotic tetracycline hydrochloride (SIGMA) at a ratio of 50 mg/ml [1] and were grown in laboratory conditions. It was continued at two temperatures of 20 and 25 °C. To eliminate and prevent the possible effects of antibiotics, all experiments were carried out in generations four and later.
Functional response test
To evaluate the functional response of Trichogramma brassicae bee populations, concentrations of 2, 5, 10, 20, 50, 65, and 85 fresh eggs of Euphtheria that were sterilized by UV rays were randomly distributed and scattered on a white cardboard strip (10 × 40 mm) were glued. For each density, female bees with a lifespan of less than 24 hours (in the case of the bisexual population, mated bees were used) in 18 replicates separately and separately for each population in test tubes (16 × 100 mm) were placed containing a certain density of host eggs. The samples were simultaneously placed in a constant temperature room (25 ± 1 °C, 60 ± 10% relative humidity, 16 hours of light, and 8 hours of darkness). After 24 hours, the whole bee insects were removed from the tubes and the samples were kept in the same environmental conditions until the parasitized eggs turned black, and finally, the number of parasitized eggs per density was recorded for each population.
The analysis of functional responses consisted of two separate steps [19, 20]. In the first step, the shape of the curve or the type of reaction function of logistic regression of the ratio of parasite eggs (Na) to the initial density of host eggs (N0) was determined based on the polynomial equation. Then, P0-P3 parameters were estimated with the CATMOD procedure in SAS software [21]. In the second step, using non-linear least square regression (NLIN procedure with DUD technique in SAS software), search power parameters or attack coefficient (a) and acquisition time (Th) were estimated. To compare the parameters estimated in different populations, a pairwise comparison was done [21].
The maximum attack rate was also calculated based on the ratio of the total test time to the time of reaching a host [22]. In many researches related to the functional response, the parameters of the attack coefficient and the acquisition time have a more comparative aspect, which in this research was also used to evaluate the effect of Wolbachia bacteria on the parasitism behavior of bisexual bees (B and CB) and unisexual bees (A) of Trichogramma brassicae bees populations were compared two by two.
Determining the exit percentage of bees stored in the cold
To evaluate the effect of the storage period on the output of each of the A, B, and CB lines grown at two temperatures of 20 and 25 °C, paper strips containing fresh Ephastia eggs were provided to each of them separately for 24 hours. Then parasitized eggs were kept in 20 glass test tubes with dimensions of 16 × 100 mm under the conditions of a bee breeding environment. After reaching the stage of pre-pupae or white-eyed pupa (approximately 10 days at 20 °C and 5 days at 25 °C), the tubes containing eggs are kept in a cold room (8 °C and total darkness) for 1, 2, 4, 5, 6, and 7 weeks were placed. After the storage period, the tubes were taken out of the cold room and transferred to a temperature of 25 °C. After the departure of the bees, while checking the sex ratio, the departure percentage of each of the A, B, and CB populations was determined and recorded.
Statistical test
For the statistical comparison of the results, after transforming the data into the square root inverse sine method, the factorial test was used based on a completely random design in the SAS software (version 6.12). Finally, the averages were compared with each other by Tukey's test.
RESULTS AND DISCUSSION
Identification of parasitoid wasps and symbiotic bacteria
In addition to the identification of the parasitoid wasp with the scientific name. Trichogramma brassicae Bezd, its bacterial contamination was also confirmed. The sequences related to the wsp gene of the Wolbachia bacterium and the ITS2 region of the Trichogramma brassicae bee infected with this bacterium have been recorded.
Functional response test
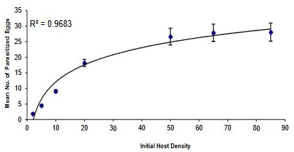
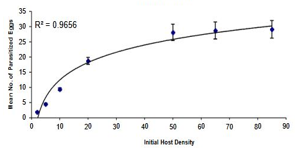
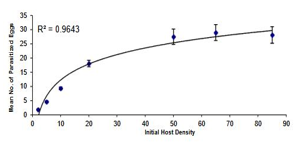
The results of logistic regression and the estimated values for the three parts of the curve of the proportion of eggs parasitized by each of the Trichogramma brassicae populations are reflected in Table 1. Given that the estimated numbers for the linear part of the curve are negative, the functional response of all three populations was determined to be of type II. The functional response curve of the bisexual (B) and bacterially infected (A) and bisexual antibiotic-treated (CB) populations to the increase in host egg density is presented in Figure 1 and the degree of data fit to the model (r2 or coefficient of determination) is presented in Table 2, which shows that the model is well fitted to the data. The search power (a) and access time (Th) parameters for the bees of the three populations A, B, and CB were obtained as 0.2393 h-1, 0.7862 h-1, 0.3016 h-1, 0.7684 h-1, 0.2525 h-1, and 0.7701 h-1, respectively. As is clear from this table, zero is located in the 95% confidence interval of the Da and DTh parameters, which indicates that the treatments do not differ significantly from each other in terms of search power and access time.
Table 1. Estimated values by logistic regression in different parts of the functional response curve of bacterially infected (A), naturally bisexual (B), and antibiotic-treated (CB) female populations of the bee Trichogramma brassicae to increasing Ephestia kuehniella egg density.
|
Line/ Population |
Coefficient |
Estimate |
SE |
χ2 |
P-value |
|
A |
P0 (Constant) |
2.9332 |
0.3557 |
68 |
< 0.001 |
|
P1 (Linear) |
-0.0449 |
0.0290 |
2.39 |
0.1218 |
|
|
P2 (Quadratic) |
-0.00047 |
0.000647 |
0.52 |
0.4689 |
|
|
P3 (Cubic) |
5.83E-6 |
4.178E-6 |
1.95 |
0.1629 |
|
|
B |
P0 (Constant) |
3.2384 |
0.3938 |
67.64 |
< 0.001 |
|
P1 (Linear) |
-0.0426 |
0.0311 |
1.88 |
0.1702 |
|
|
P2 (Quadratic) |
-0.00067 |
0.000684 |
0.96 |
0.3276 |
|
|
P3 (Cubic) |
7.47E-6 |
4.39E-6 |
2.90 |
0.0888 |
|
|
CB |
P0 (Constant) |
3.5689 |
0.4047 |
77.77 |
< 0.001 |
|
P1 (Linear) |
-0.0882 |
0.0317 |
7.74 |
0.0054 |
|
|
P2 (Quadratic) |
0.000449 |
0.000693 |
0.42 |
0.5175 |
|
|
P3 (Cubic) |
-1.22E-8 |
4.429E-6 |
0.00 |
0.9978 |
|
|
|
|
a) |
b) |
|
|
|
|
c) |
|
|
Figure 1. The functional response curve of bacterially infected, a) naturally bisexual, b) and antibiotic-treated, c) female populations of the bee Trichogramma brassicae to changes in Ephestia kuehniella egg density. The bars and curves represent the observed mean (mean ± SE) and the model-predicted values, respectively. |
|
Table 2. Functional response parameters of bacterially infected (A), naturally bisexual (B), and antibiotic-treated (CB) populations of Trichogramma brassicae to different densities of Ephestia kehniella eggs, estimated by the Disk equation.
|
Line/Population |
Type |
r2 |
Parameter |
Estimate (Mean ± SE) |
Asymptotic 95% CI* |
|
|
Lower |
Upper |
|||||
|
A |
II |
0.968 |
a |
0.2393 ± 0.0717 |
0.0973 |
0.3813 |
|
Th |
0.7862 ± 0.0323 |
0.7223 |
0.8501 |
|||
|
T / Th |
30.52 |
- |
- |
|||
|
B |
II |
0.966 |
a |
0.3016 ± 0.1263 |
0.0515 |
0.5516 |
|
Th |
0.7684 ± 0.0351 |
0.6989 |
0.8378 |
|||
|
T / Th |
31.23 |
- |
- |
|||
|
CB |
II |
0.964 |
a |
0.2525 ± 0.0831 |
0.0881 |
0.4170 |
|
Th |
0.7701 ± 0.0332 |
0.7044 |
0.8358 |
|||
|
T / Th |
31.16 |
- |
- |
|||
|
A vs B |
- |
- |
Da |
0.0622 ± 0.1419 |
-0.3417 |
0.2172 |
|
DTh |
0.0178 ± 0.0478 |
-0.0763 |
0.1119 |
|||
|
A vs CB |
- |
- |
Da |
-0.0132 ± 0.1095 |
-0.2289 |
0.2025 |
|
DTh |
0.0161 ± 0.0463 |
-0.0752 |
0.1073 |
|||
|
B vs CB |
- |
- |
Da |
0.0490 ± 0.1495 |
-0.2454 |
0.3435 |
|
DTh |
-0.00174 ± 0.0483 |
-0.0969 |
0.0935 |
|||
Percentage of exit of bees stored in cold
Analysis of variance of the total data, which was a combination of three factors of bee population (three populations), rearing temperature (20 and 25 °C), and storage time (1-7 weeks), showed that the treatments had a significant difference at the one percent probability level (P = 0.0001, F41,168 = 41.34). Also, the difference between the treatments related to each of the factors was significant and there was an interaction between them (P = 0.0001). According to the results obtained in this study, the percentage of the exodus of bees infected with bacteria (A), naturally bisexual (B), and antibiotic-treated bisexual (CB) was not significantly different from each other, regardless of the effect of rearing temperature, and all three were placed in the same group (Table 3). Theoretically, the highest parasitism or maximum attack rate by female-producing unisexual, naturally bisexual, and antibiotic-treated bisexual populations was determined to be 30.52, 31.23, and 31.16 host eggs per day, respectively, and the numerical value of this index in the naturally bisexual population B was higher than that of the two populations CB and A.
Table 3. Percentage of hatching of parasitized Ephestia kuehniella eggs by bacterially infected (A), naturally bisexual (B), and antibiotic-treated (CB) female populations of Trichogramma brassicae after storage under cold storage conditions (Mean ± SE).
|
Population (Rearing temperature) |
Storage time (day) |
Total Mean |
||||||
|
7 |
14 |
21 |
28 |
35 |
42 |
49 |
||
|
A (20°C) |
87.76 ± 0.68a |
85.1 ± 0.71ab |
86.72 ± 0.33ab |
78.38 ± 1.3ab |
75.52 ± 2.46bc |
58.16 ± 6.24cd |
49.74 ± 3.05cde |
74.48A |
|
A (25°C) |
88.62 ± 1.22a |
74.72 ± 4.03bc |
46.54 ± 2.82de |
42.72 ± 4.18def |
40.6 ± 4.78def |
35.34 ± 3.74ef |
14.86 ± 1.16g |
49.05B |
|
B (20°C) |
90.42 ± 2.2a |
89.26 ± 1.44a |
88.56 ± 1.8a |
77.74 ± 1.74ab |
74.4 ± 3.57bc |
49.7 ± 6.25cde |
47.14 ± 3.98de |
73.89A |
|
B (25°C) |
88.20 ± 1.57a |
67.76 ± 5.81c |
52.94 ± 5.19de |
47.74 ± 5.35de |
43.28 ± 4.47def |
41.04 ± 1.65def |
34.1 ± 3.71ef |
53.58B |
|
CB (20°C) |
87.12 ± 1.03ab |
85.32 ± 0.97ab |
88.46 ± 1.08a |
76 ± 2.75bc |
74.94 ± 2.9bc |
57.52 ± 3.87cd |
49.22 ± 3.87cde |
74.14A |
|
CB (25°C) |
88.82 ± 1.37a |
66.28 ± 5.7cd |
48.24 ± 2.2cde |
41.52 ± 5.29ef |
35.52 ± 2.53ef |
32.84 ± 4.33ef |
27.14 ± 1.85f |
48.62B |
|
Total mean |
84.4A |
78.07B |
68.57C |
60.75D |
57.37E |
45.7F |
37.03G |
- |
The results also show that in all three populations, the temperature at which the bees were reared before storage played a very important role in the exodus rate, and especially from the third week onwards, their differences were significant and they were distinguished from each other in terms of grouping. Overall, after 49 days of storage, among the treatments, population A (infected with Wolbachia) grown at 20 °C had the highest numerical percentage of emergence (49.74 ± 3.05), and conversely, population A grown at 25 °C had the lowest percentage of emergence (14.68 ± 1.16) (Table 3).
As in most previous reports on Trichogramma bees in vitro, a functional type II response was obtained for all three populations infected and uninfected with Trichogramma brassicae. Some studies on native populations of Trichogramma brassicae at 25 °C have shown a type II response to grain moth eggs [23]. Before this study, the functional response of Trichogramma bees infected with Wolbachia bacteria to the second type of cereal weevil (Sitotroga cerealella live) was determined [24].
In this study, the access time and search power of the female population and the bisexual population did not show a significant difference. However, in the study [24], the access time of the female population was estimated to be significantly longer than that of the natural bisexual population, which was attributed to the difference in the infected bee population. In this study, as mentioned, the maximum attack rate by the female monosexual (A) and natural bisexual (B) populations was determined to be 30.52 and 31.23, respectively, which shows a significant difference from the results of Farrokhi et al. [24]. The reason for the difference is the difference in the two intermediate host species used in this study. Overall, the differences in these parameters estimated for a single insect species can be influenced by various factors such as plant, host, temperature, and type of prey or host [25, 26].
The storage of Trichogramma is of great importance in the development of applications of this egg parasitoid wasp. Fortunately, many studies have been conducted on short-term and long-term storage of Trichogramma bees in cold storage. For different species, it is possible to store bees in the egg, larva, pre-pupa, and pupa stages for 2 weeks to 4 months at temperatures of 2-13 degrees, in a simple (one-stage) or with diapause induction (two-stage) [27, 28].
The results of a study conducted by Shi et al. [29] on the effect of diapause induction and long-term storage of two ecotypes of the bee Trichogramma dendrolimi Matsumura showed the effect of environmental conditions on the sensitivity of bees to cold. In such a way the ecotype from the cold regions of northern China showed less sensitivity and higher reproductive power compared to the ecotype collected from southern China. In this study, although Wolbachia bacteria increased after storage of bees Trichogramma brassicae reared at 25 °C reduced their hatching percentage, but if the female bees were reared for several generations at 20 °C before storage, not only was there no negative effect on the hatching percentage, but the numerical average of the hatching percentage of female bees of line (A) after 49 days was higher than that of bisexual populations (B) and (CB) (Table 3), which could indicate an increase in cold tolerance due to an increase in the concentration and density of Wolbachia bacteria in the host insect body. The results obtained by Farrokhi et al. [24] in this regard are completely consistent and similar to the results of this study.
So far, studies conducted on the effect of Wolbachia bacteria on the hatching percentage of infected bees stored at low temperatures or that have entered diapause have not only shown no adverse or harmful effects, but some reports contain positive points in terms of practicality and the possibility of storing unisexual populations. According to Wang and Smith [30], female populations of Trichogramma mintum Riley can tolerate low temperatures better than bisexual populations. These populations also showed a lower thermal threshold for growth and development and their percentage of emergence after a storage period at 4 °C was somewhat higher. Pintureau et al. [31] also investigated the side effects of this bacterium on the diapause of Trichogramma evanescens Westwood, which was collected from a warm Mediterranean climate, and did not observe any positive or negative effects on the emergence rate of stored bees or their fertility rate. Finally, they considered the use of unisexual populations infected with the bacteria to be advisable, especially in terms of increasing production efficiency and reducing costs. From these results, it can be concluded that this symbiotic bacterium is capable of increasing the survival of the host during the cold season.
Despite some opinions that Wolbachia bacteria should be removed from Trichogramma colony cultivation due to some negative effects on the biological indicators of the host bee [28], the results of this study and previous studies showed that despite some negative effects of the bacteria on their host, no statistical difference was observed between the female-bearing populations infected with the bacteria and the hermaphrodite populations in terms of distribution, foraging, parasitism, fertility, and lifespan. In practical terms, the results indicate that the female-bearing form is more efficient in greenhouse and field conditions [32].
Recently Ebrahimi et al. [33] have conducted studies and suggested the selection of a unisexual recombinant line of the bee Trichogramma pretiosum Riley infected with Wolbachia bacteria for use in applied biological control programs. What can be suggested here is the need for further investigation into finding new infected species in different climatic conditions, especially at high altitudes, where there are more suitable conditions for the stabilization of the bacteria in their host. Also, the investigation of the horizontal transfer of Wolbachia bacteria to improve the quality of strains of Trichogramma that have special capabilities (such as less sensitivity to dryness and heat or high flight and search power) and comparing its biological indicators with a bisexual and uninfected population of the same ecotype, to plan with greater awareness to eliminate the possible undesirable effects of symbiotic microorganisms.
CONCLUSION
In this study, the effect of the intracellular symbiotic bacterium Wolbachia on the search power, access time, and exit percentage of female (A), natural hermaphrodite (B), and antibiotic-treated hermaphrodite (CB) Trichogramma brassicae wasps was investigated in the laboratory and cold storage. Based on the results of short-term cold storage, the average total exit percentage of natural hermaphrodite, bacterially infected female, and antibiotic-treated hermaphrodite bees reared at 20 °C was significantly higher than that of 25 °C. However, during similar storage periods, all three populations were placed in common groups in terms of exit percentage without significant differences. According to the results of this study and previous findings, by providing optimal conditions for the reproduction and maintenance of Wolbachia-infected lines, the use of unisexual bees can have a relative advantage compared to natural bisexual bees.
ACKNOWLEDGMENTS: None
CONFLICT OF INTEREST: None
FINANCIAL SUPPORT: None
ETHICS STATEMENT: None